

CaM kinase II initiates meiotic spindle depolymerization independently of APC/C activation
- PMID: 19064669
- PMCID: PMC2600749
- DOI: 10.1083/jcb.200807006
CaM kinase II initiates meiotic spindle depolymerization independently of APC/C activation
Abstract
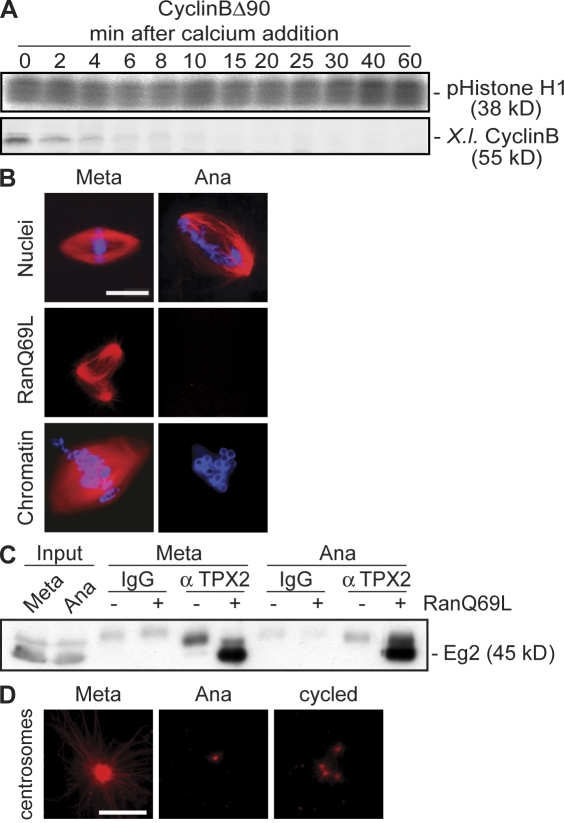
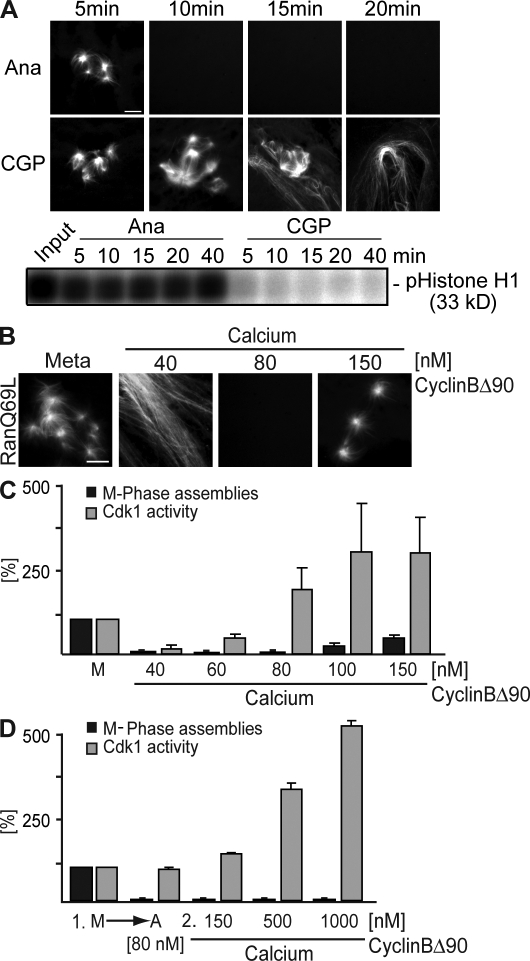
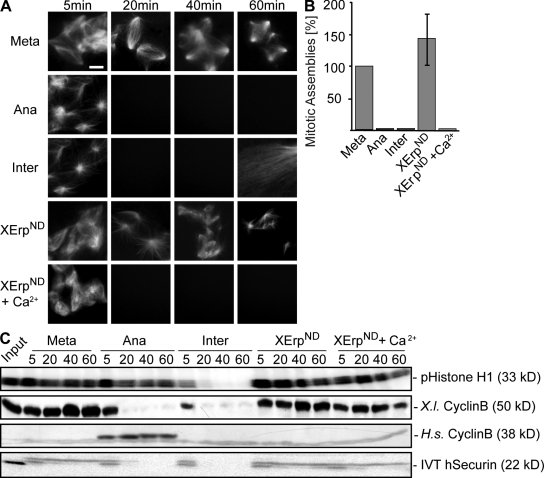
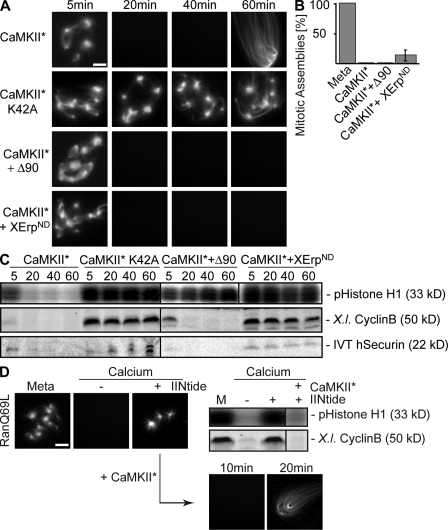
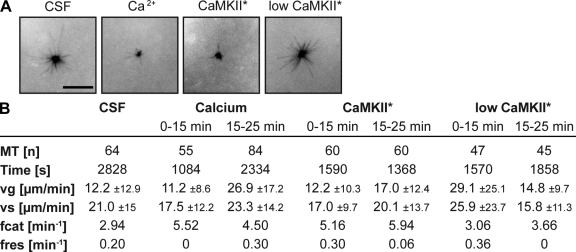
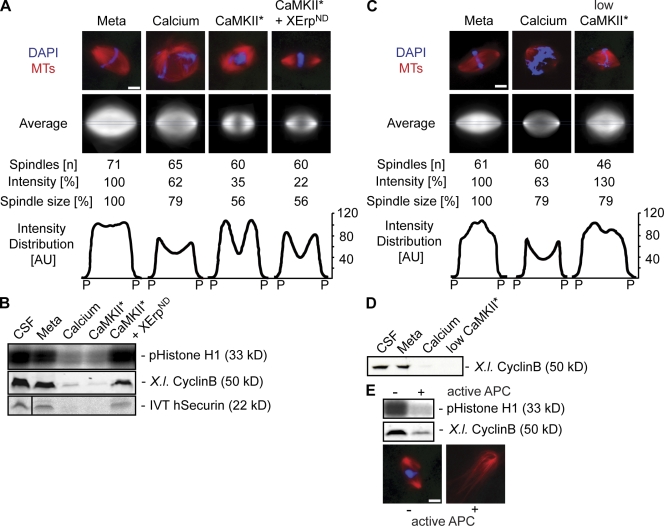
Altered spindle microtubule dynamics at anaphase onset are the basis for chromosome segregation. In Xenopus laevis egg extracts, increasing free calcium levels and subsequently rising calcium-calmodulin-dependent kinase II (CaMKII) activity promote a release from meiosis II arrest and reentry into anaphase. CaMKII induces the activation of the anaphase-promoting complex/cyclosome (APC/C), which destines securin and cyclin B for degradation to allow chromosome separation and mitotic exit. In this study, we investigated the calcium-dependent signal responsible for microtubule depolymerization at anaphase onset after release from meiotic arrest in Xenopus egg extracts. Using Ran-guanosine triphosphate-mediated microtubule assemblies and quantitative analysis of complete spindles, we demonstrate that CaMKII triggers anaphase microtubule depolymerization. A CaMKII-induced twofold increase in microtubule catastrophe rates can explain reduced microtubule stability. However, calcium or constitutively active CaMKII promotes microtubule destabilization even upon APC/C inhibition and in the presence of high cyclin-dependent kinase 1 activity. Therefore, our data demonstrate that CaMKII turns on parallel pathways to activate the APC/C and to induce microtubule depolymerization at meiotic anaphase onset.
Figures
Similar articles
-
Calcium triggers exit from meiosis II by targeting the APC/C inhibitor XErp1 for degradation.Nature. 2005 Oct 13;437(7061):1048-52. doi: 10.1038/nature04093. Epub 2005 Aug 28. Nature. 2005. PMID: 16127448
-
Calcium elevation at fertilization coordinates phosphorylation of XErp1/Emi2 by Plx1 and CaMK II to release metaphase arrest by cytostatic factor.Curr Biol. 2005 Aug 23;15(16):1458-68. doi: 10.1016/j.cub.2005.07.030. Curr Biol. 2005. PMID: 16040245
-
Cyclin B3 activates the Anaphase-Promoting Complex/Cyclosome in meiosis and mitosis.PLoS Genet. 2020 Nov 2;16(11):e1009184. doi: 10.1371/journal.pgen.1009184. eCollection 2020 Nov. PLoS Genet. 2020. PMID: 33137813 Free PMC article.
-
Preventing fatal destruction: inhibitors of the anaphase-promoting complex in meiosis.Cell Cycle. 2006 Feb;5(4):405-15. doi: 10.4161/cc.5.4.2476. Epub 2006 Feb 15. Cell Cycle. 2006. PMID: 16479160 Review.
-
Mammalian egg activation: from Ca2+ spiking to cell cycle progression.Reproduction. 2005 Dec;130(6):813-23. doi: 10.1530/rep.1.00710. Reproduction. 2005. PMID: 16322541 Review.
Cited by
-
CMTX Disorder and CamKinase.Front Cell Neurosci. 2016 Mar 1;10:49. doi: 10.3389/fncel.2016.00049. eCollection 2016. Front Cell Neurosci. 2016. PMID: 26973463 Free PMC article. No abstract available.
-
XMAP215 activity sets spindle length by controlling the total mass of spindle microtubules.Nat Cell Biol. 2013 Sep;15(9):1116-22. doi: 10.1038/ncb2834. Epub 2013 Aug 25. Nat Cell Biol. 2013. PMID: 23974040
-
Modulation of cell cycle control during oocyte-to-embryo transitions.EMBO J. 2013 Aug 14;32(16):2191-203. doi: 10.1038/emboj.2013.164. Epub 2013 Jul 26. EMBO J. 2013. PMID: 23892458 Free PMC article. Review.
-
Cilia and cell cycle re-entry: more than a coincidence.Cell Cycle. 2011 Aug 15;10(16):2683-90. doi: 10.4161/cc.10.16.17009. Epub 2011 Aug 15. Cell Cycle. 2011. PMID: 21814045 Free PMC article. Review.
-
Calcium Oscillatory Patterns and Oocyte Activation During Fertilization: a Possible Mechanism for Total Fertilization Failure (TFF) in Human In Vitro Fertilization?Reprod Sci. 2021 Mar;28(3):639-648. doi: 10.1007/s43032-020-00293-5. Epub 2020 Aug 19. Reprod Sci. 2021. PMID: 32813196 Review.
References
-
- Blower, M.D., M. Nachury, R. Heald, and K. Weis. 2005. A Rae1-containing ribonucleoprotein complex is required for mitotic spindle assembly. Cell. 121:223–234. - PubMed
-
- Carazo-Salas, R.E., G. Guarguaglini, O.J. Gruss, A. Segref, E. Karsenti, and I.W. Mattaj. 1999. Generation of GTP-bound Ran by RCC1 is required for chromatin-induced mitotic spindle formation. Nature. 400:178–181. - PubMed
-
- Carazo-Salas, R.E., O.J. Gruss, I.W. Mattaj, and E. Karsenti. 2001. Ran-GTP coordinates regulation of microtubule nucleation and dynamics during mitotic-spindle assembly. Nat. Cell Biol. 3:228–234. - PubMed
-
- Cassimeris, L., and R.V. Skibbens. 2003. Regulated assembly of the mitotic spindle: a perspective from two ends. Curr. Issues Mol. Biol. 5:99–112. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
