

Prion switching in response to environmental stress
- PMID: 19067491
- PMCID: PMC2586387
- DOI: 10.1371/journal.pbio.0060294
Prion switching in response to environmental stress
Abstract
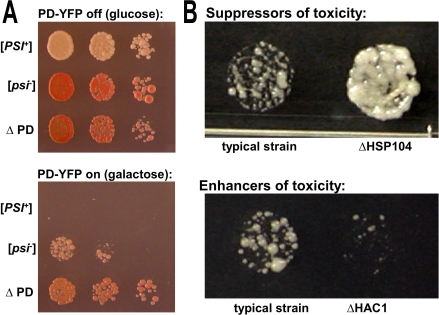
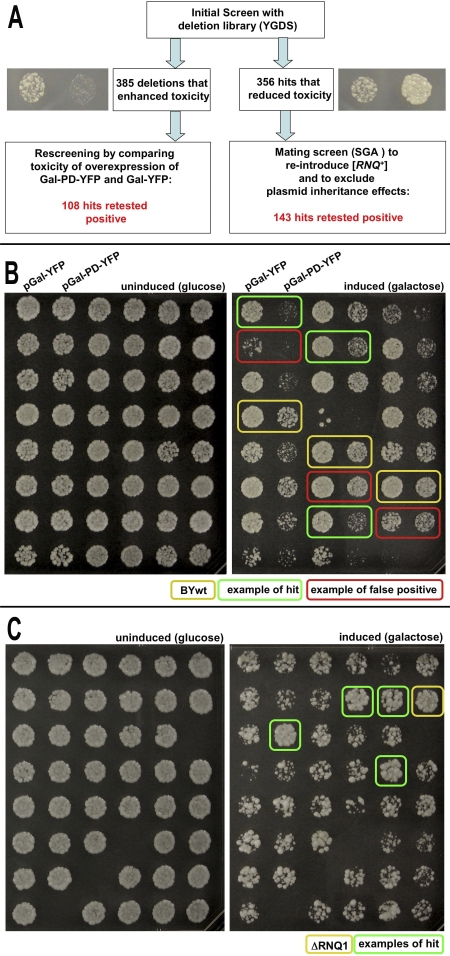
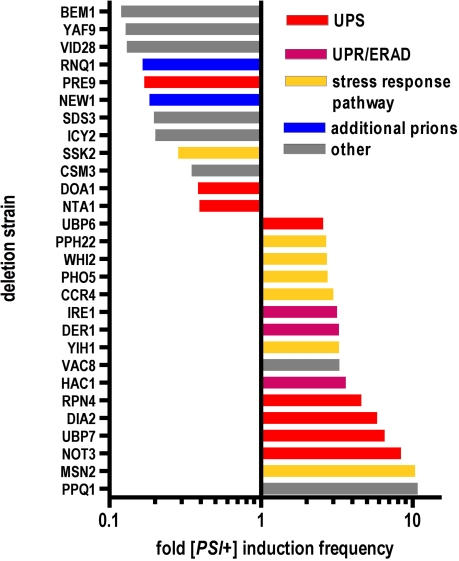
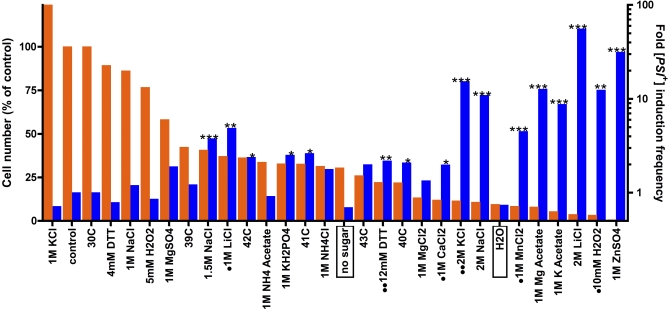
Evolution depends on the manner in which genetic variation is translated into new phenotypes. There has been much debate about whether organisms might have specific mechanisms for "evolvability," which would generate heritable phenotypic variation with adaptive value and could act to enhance the rate of evolution. Capacitor systems, which allow the accumulation of cryptic genetic variation and release it under stressful conditions, might provide such a mechanism. In yeast, the prion [PSI(+)] exposes a large array of previously hidden genetic variation, and the phenotypes it thereby produces are advantageous roughly 25% of the time. The notion that [PSI(+)] is a mechanism for evolvability would be strengthened if the frequency of its appearance increased with stress. That is, a system that mediates even the haphazard appearance of new phenotypes, which have a reasonable chance of adaptive value would be beneficial if it were deployed at times when the organism is not well adapted to its environment. In an unbiased, high-throughput, genome-wide screen for factors that modify the frequency of [PSI(+)] induction, signal transducers and stress response genes were particularly prominent. Furthermore, prion induction increased by as much as 60-fold when cells were exposed to various stressful conditions, such as oxidative stress (H2O2) or high salt concentrations. The severity of stress and the frequency of [PSI(+)] induction were highly correlated. These findings support the hypothesis that [PSI(+)] is a mechanism to increase survival in fluctuating environments and might function as a capacitor to promote evolvability.
Conflict of interest statement
Figures

References
-
- Cox B. [PSI], a cytoplasmic suppressor of super-suppression in yeast. Heredity. 1965;20:505–521.
-
- Tuite MF, Cox BS. Propagation of yeast prions. Nat Rev Mol Cell Biol. 2003;4:878–890. - PubMed
-
- Shorter J, Lindquist S. Prions as adaptive conduits of memory and inheritance. Nat Rev Genet. 2005;6:435–450. - PubMed
-
- Chien P, Weissman JS, DePace AH. Emerging principles of conformation-based prion inheritance. Annu Rev Biochem. 2004;73:617–656. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
