

Nongenomic steroid-triggered oocyte maturation: of mice and frogs
- PMID: 19071151
- PMCID: PMC2702721
- DOI: 10.1016/j.steroids.2008.11.010
Nongenomic steroid-triggered oocyte maturation: of mice and frogs
Abstract
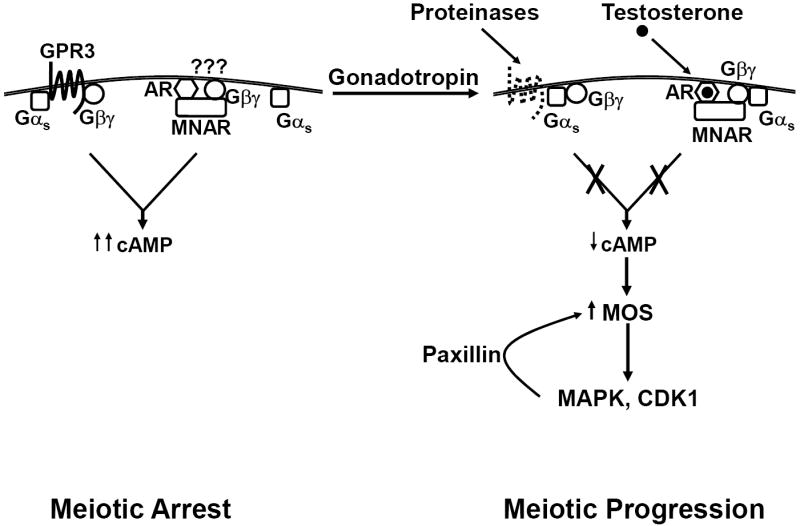
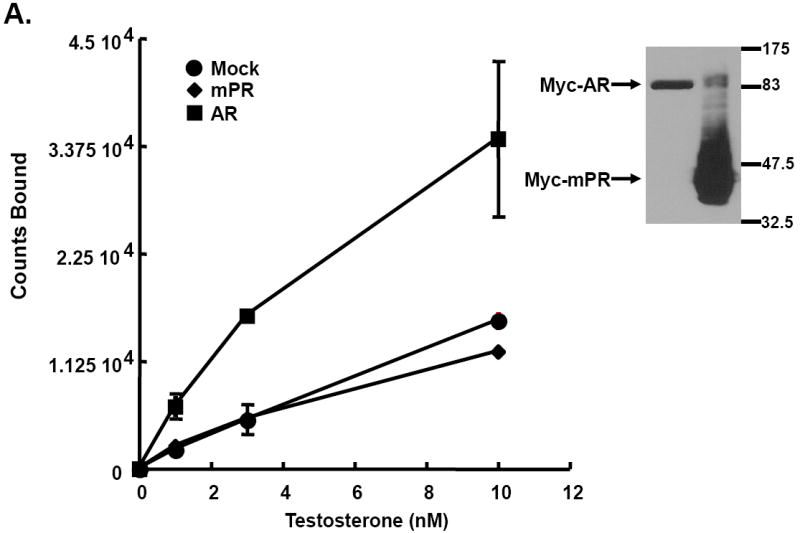
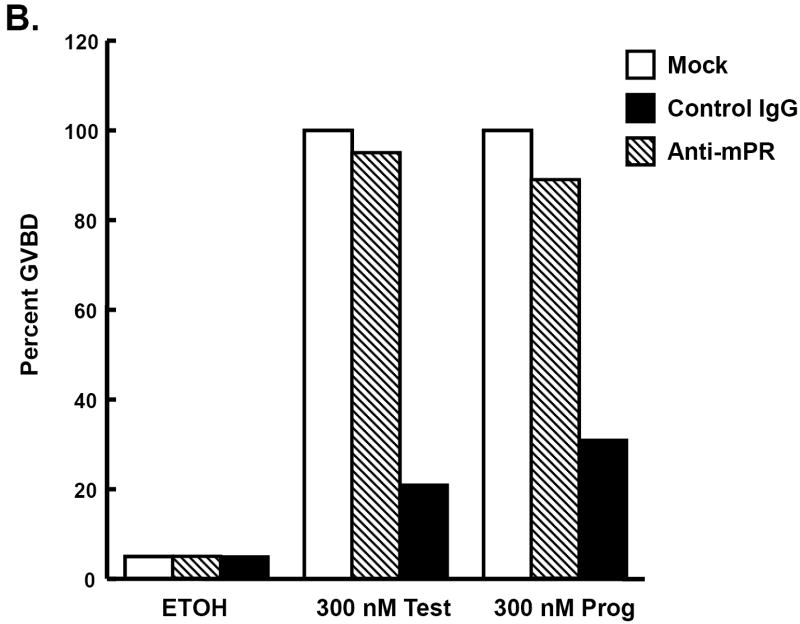
Luteinizing hormone (LH) mediates many important processes in ovarian follicles, including cumulus cell expansion, changes in gap junction expression and activity, sterol and steroid production, and the release of paracrine signaling molecules. All of these functions work together to trigger oocyte maturation (meiotic progression) and subsequent ovulation. Many laboratories are interested in better understanding both the extra-oocyte follicular processes that trigger oocyte maturation, as well as the intra-oocyte molecules and signals that regulate meiosis. Multiple model systems have been used to study LH-effects in the ovary, including fish, frogs, mice, rats, pigs, and primates. Here we provide a brief summary of oocyte maturation, focusing primarily on steroid-triggered meiotic progression in frogs and mice. Furthermore, we present new studies that implicate classical steroid receptors rather than alternative non-classical membrane steroid receptors as the primary regulators of steroid-mediated oocyte maturation in both of these model systems.
Figures

Similar articles
-
Specific modulation of nongenomic androgen signaling in the ovary.Steroids. 2005 May-Jun;70(5-7):352-60. doi: 10.1016/j.steroids.2005.02.008. Epub 2005 Mar 16. Steroids. 2005. PMID: 15862817
-
Epidermal growth factor receptor signaling is required for normal ovarian steroidogenesis and oocyte maturation.Proc Natl Acad Sci U S A. 2005 Nov 8;102(45):16257-62. doi: 10.1073/pnas.0508521102. Epub 2005 Oct 31. Proc Natl Acad Sci U S A. 2005. PMID: 16260720 Free PMC article.
-
The physiology of the Xenopus laevis ovary.Methods Mol Biol. 2006;322:17-30. doi: 10.1007/978-1-59745-000-3_2. Methods Mol Biol. 2006. PMID: 16739713 Review.
-
Steroids and oocyte maturation--a new look at an old story.Mol Endocrinol. 2004 Apr;18(4):769-75. doi: 10.1210/me.2003-0317. Epub 2003 Nov 20. Mol Endocrinol. 2004. PMID: 14630999 Review.
-
Luteinizing Hormone Action in Human Oocyte Maturation and Quality: Signaling Pathways, Regulation, and Clinical Impact.Reprod Sci. 2020 Jun;27(6):1223-1252. doi: 10.1007/s43032-019-00137-x. Epub 2020 Jan 6. Reprod Sci. 2020. PMID: 32046451 Free PMC article. Review.
Cited by
-
Protein-tyrosine kinase signaling in the biological functions associated with sperm.J Signal Transduct. 2012;2012:181560. doi: 10.1155/2012/181560. Epub 2012 Nov 11. J Signal Transduct. 2012. PMID: 23209895 Free PMC article.
-
Understanding extranuclear (nongenomic) androgen signaling: what a frog oocyte can tell us about human biology.Steroids. 2011 Aug;76(9):822-8. doi: 10.1016/j.steroids.2011.02.016. Epub 2011 Feb 25. Steroids. 2011. PMID: 21354434 Free PMC article. Review.
-
What Do We Know about Classical and Non-Classical Progesterone Receptors in the Human Female Reproductive Tract? A Review.Int J Mol Sci. 2021 Oct 19;22(20):11278. doi: 10.3390/ijms222011278. Int J Mol Sci. 2021. PMID: 34681937 Free PMC article. Review.
-
Roles of progesterone receptor membrane component 1 and membrane progestin receptor alpha in regulation of zebrafish oocyte maturation.Gen Comp Endocrinol. 2018 Jul 1;263:51-61. doi: 10.1016/j.ygcen.2018.04.009. Epub 2018 Apr 9. Gen Comp Endocrinol. 2018. PMID: 29649418 Free PMC article.
-
Overlapping nongenomic and genomic actions of thyroid hormone and steroids.Best Pract Res Clin Endocrinol Metab. 2015 Aug;29(4):581-93. doi: 10.1016/j.beem.2015.04.001. Epub 2015 Apr 22. Best Pract Res Clin Endocrinol Metab. 2015. PMID: 26303085 Free PMC article. Review.
References
-
- Albertini DF, Carabatsos MJ. Comparative aspects of meiotic cell cycle control in mammals. J Mol Med. 1998;76(12):795–9. - PubMed
-
- Hammes SR. Steroids and oocyte maturation--a new look at an old story. Mol Endocrinol. 2004;18(4):769–75. - PubMed
-
- Maller JL, Krebs EG. Regulation of oocyte maturation. Curr Top Cell Regul. 1980;16:271–311. - PubMed
-
- Rasar MA, Hammes SR. The physiology of the Xenopus laevis ovary. Methods Mol Biol. 2006;322:17–30. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
