

Age-dependent signature of metallothionein expression in primary CD4 T cell responses is due to sustained zinc signaling
- PMID: 19072254
- PMCID: PMC2848531
- DOI: 10.1089/rej.2008.0747
Age-dependent signature of metallothionein expression in primary CD4 T cell responses is due to sustained zinc signaling
Abstract
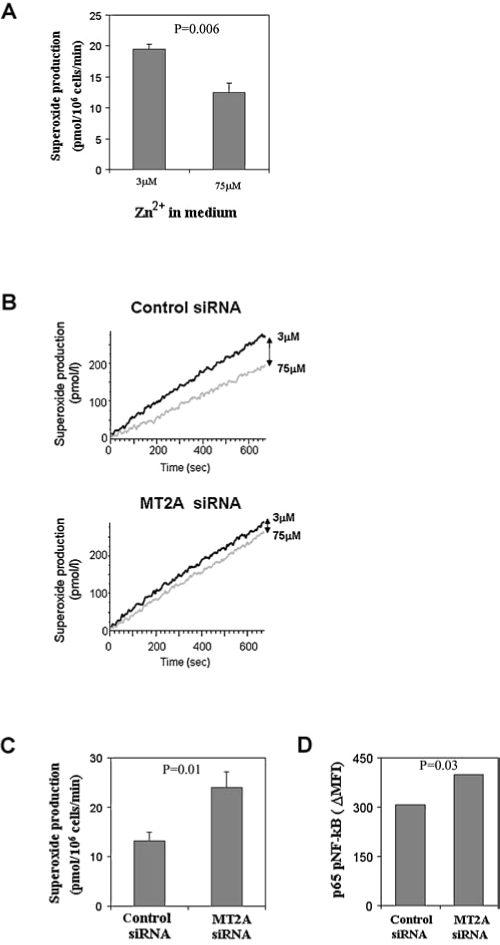
The ability to mount adaptive immune responses to vaccinations and viral infections declines with increasing age. To identify mechanisms leading to immunosenescence, primary CD4 T cell responses were examined in 60- to 75-year-old individuals lacking overt functional defects. Transcriptome analysis indicated a selective defect in zinc homeostasis. CD4 T cell activation was associated with zinc influx via the zinc transporter Zip6, leading to increased free cytoplasmic zinc and activation of negative feedback loops, including the induction of zinc-binding metallothioneins. In young adults, activation-induced cytoplasmic zinc concentrations declined after 2 days to below prestimulation levels. In contrast, activated naïve CD4 T cells from older individuals failed to downregulate cytoplasmic zinc, resulting in excessive induction of metallothioneins. Activation-induced metallothioneins regulated the redox state in activated T cells and accounted for an increased proliferation of old CD4 T cells, suggesting that regulation of T cell zinc homeostasis functions as a compensatory mechanism to preserve the replicative potential of naïve CD4 T cells with age.
Figures






Similar articles
-
Metallothionein regulates intracellular zinc signaling during CD4(+) T cell activation.BMC Immunol. 2016 Jun 2;17(1):13. doi: 10.1186/s12865-016-0151-2. BMC Immunol. 2016. PMID: 27251638 Free PMC article.
-
Zap70 Regulates TCR-Mediated Zip6 Activation at the Immunological Synapse.Front Immunol. 2021 Jul 29;12:687367. doi: 10.3389/fimmu.2021.687367. eCollection 2021. Front Immunol. 2021. PMID: 34394081 Free PMC article.
-
Metallothioneins/PARP-1/IL-6 interplay on natural killer cell activity in elderly: parallelism with nonagenarians and old infected humans. Effect of zinc supply.Mech Ageing Dev. 2003 Apr;124(4):459-68. doi: 10.1016/s0047-6374(03)00023-x. Mech Ageing Dev. 2003. PMID: 12714254
-
NK and NKT cells in aging and longevity: role of zinc and metallothioneins.J Clin Immunol. 2009 Jul;29(4):416-25. doi: 10.1007/s10875-009-9298-4. Epub 2009 May 1. J Clin Immunol. 2009. PMID: 19408107 Review.
-
Zinc, metallothioneins and immunosenescence.Proc Nutr Soc. 2010 Aug;69(3):290-9. doi: 10.1017/S0029665110001862. Epub 2010 Jun 28. Proc Nutr Soc. 2010. PMID: 20579408 Review.
Cited by
-
Regulatory Role of Zinc in Immune Cell Signaling.Mol Cells. 2021 May 31;44(5):335-341. doi: 10.14348/molcells.2021.0061. Mol Cells. 2021. PMID: 33986184 Free PMC article. Review.
-
Zinc as a Gatekeeper of Immune Function.Nutrients. 2017 Nov 25;9(12):1286. doi: 10.3390/nu9121286. Nutrients. 2017. PMID: 29186856 Free PMC article. Review.
-
Immunosenescence, aging, and systemic lupus erythematous.Autoimmune Dis. 2013;2013:267078. doi: 10.1155/2013/267078. Epub 2013 Oct 24. Autoimmune Dis. 2013. PMID: 24260712 Free PMC article. Review.
-
Zinc Ionophore Pyrithione Mimics CD28 Costimulatory Signal in CD3 Activated T Cells.Int J Mol Sci. 2024 Apr 12;25(8):4302. doi: 10.3390/ijms25084302. Int J Mol Sci. 2024. PMID: 38673887 Free PMC article.
-
Signal inhibition by the dual-specific phosphatase 4 impairs T cell-dependent B-cell responses with age.Proc Natl Acad Sci U S A. 2012 Apr 10;109(15):E879-88. doi: 10.1073/pnas.1109797109. Epub 2012 Mar 20. Proc Natl Acad Sci U S A. 2012. PMID: 22434910 Free PMC article.
References
-
- Thompson WW. Shay DK. Weintraub E. Brammer L. Cox N. Anderson LJ. Fukuda K. Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA. 2003;289:179–186. - PubMed
-
- Gavazzi G. Krause KH. Ageing and infection. Lancet Infect Dis. 2002;2:659–666. - PubMed
-
- Donahue JG. Choo PW. Manson JE. Platt R. The incidence of herpes zoster. Arch Intern Med. 1995;155:1605–1609. - PubMed
-
- Grubeck-Loebenstein B. Wick G. The aging of the immune system. Adv Immunol. 2002;80:243–284. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
