

RhoA downstream of G(q) and G(12/13) pathways regulates protease-activated receptor-mediated dense granule release in platelets
- PMID: 19073150
- PMCID: PMC2745276
- DOI: 10.1016/j.bcp.2008.11.017
RhoA downstream of G(q) and G(12/13) pathways regulates protease-activated receptor-mediated dense granule release in platelets
Abstract
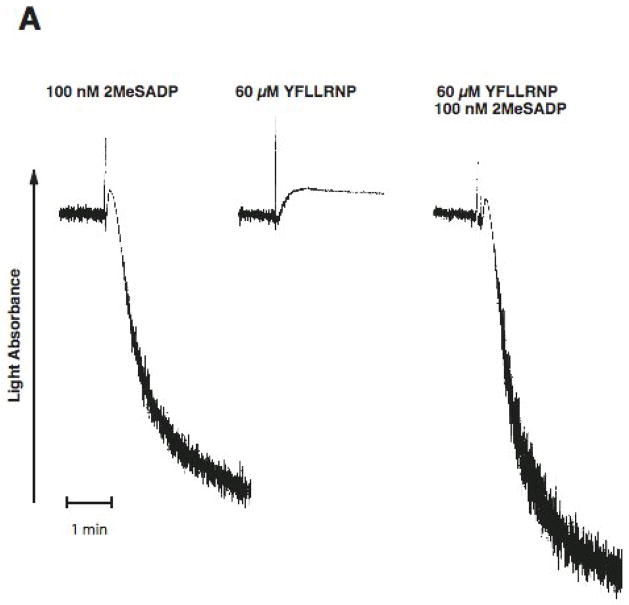
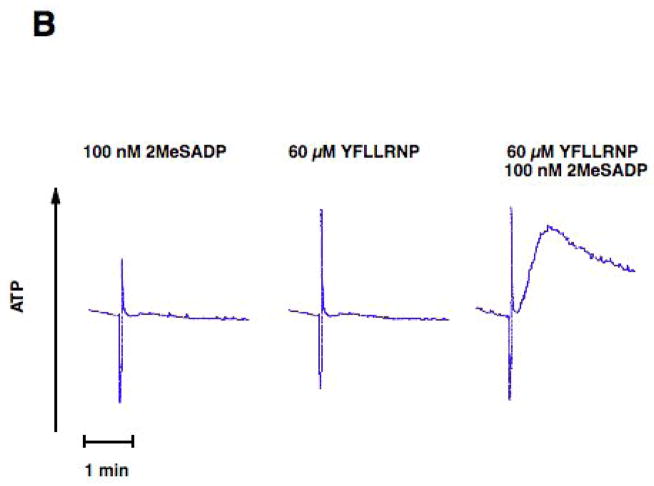
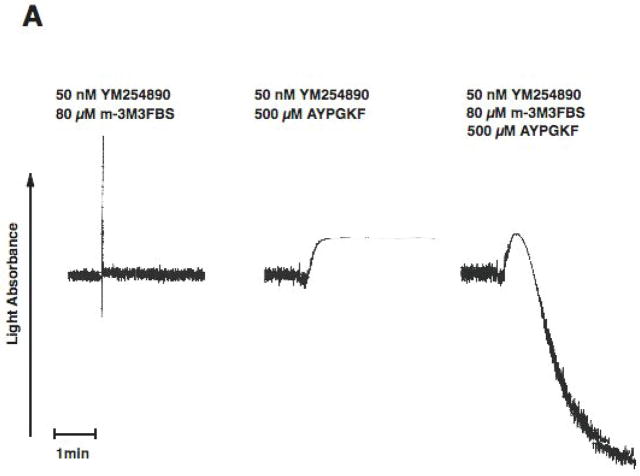
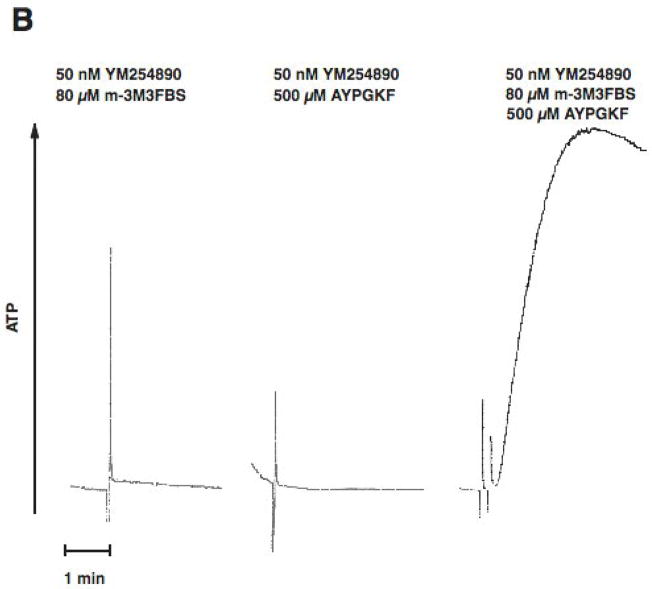
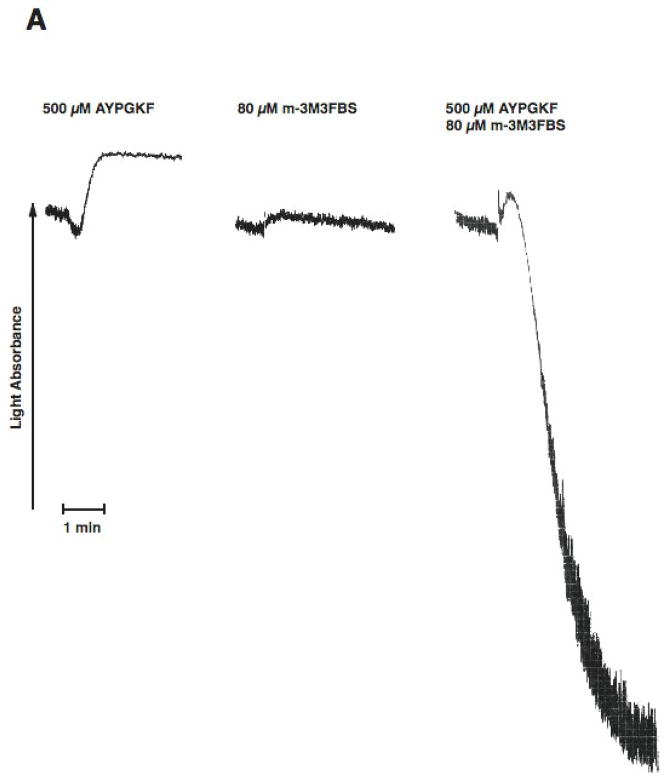
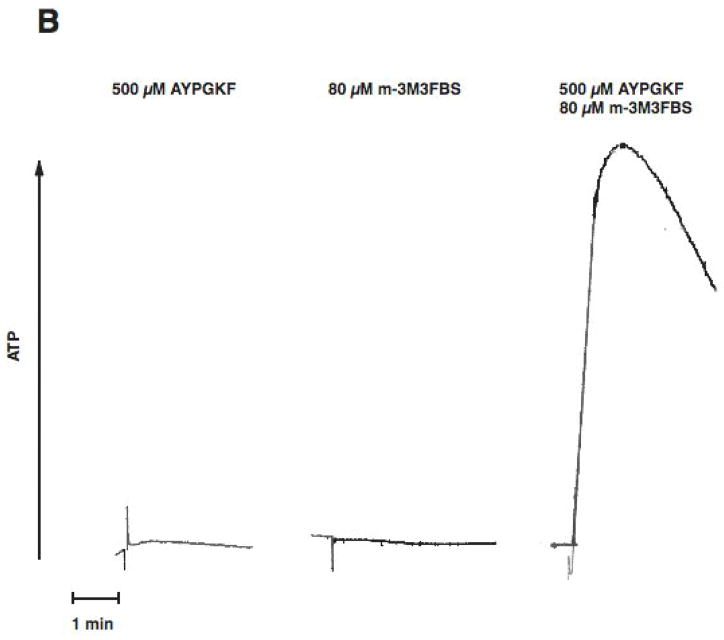
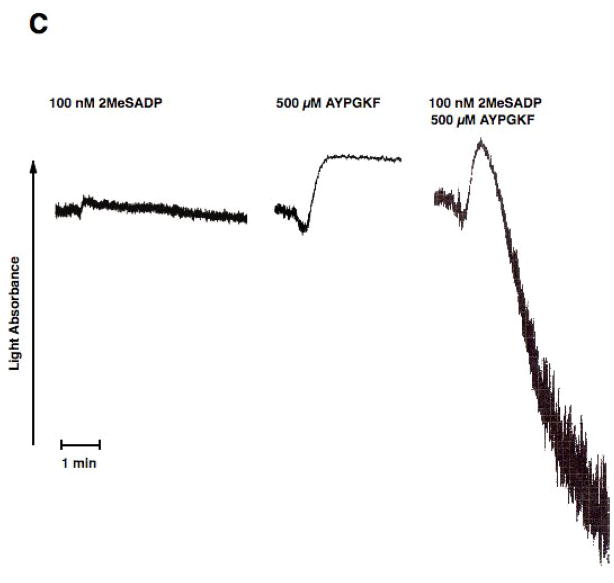
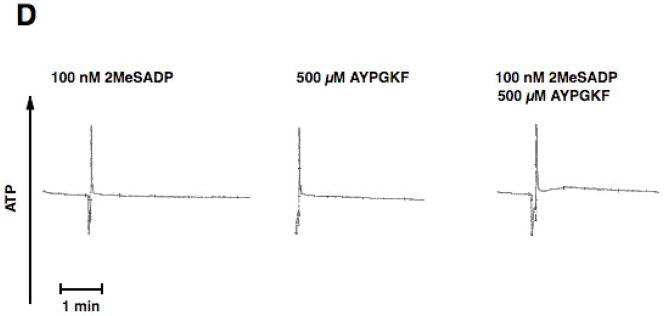
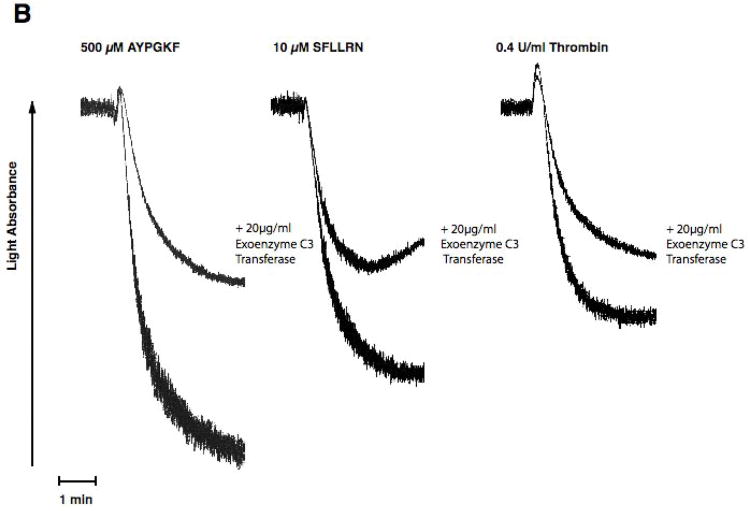
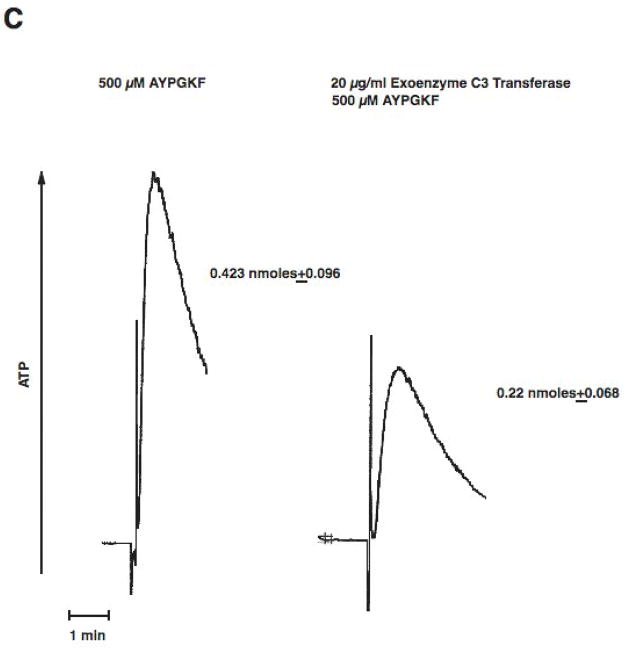
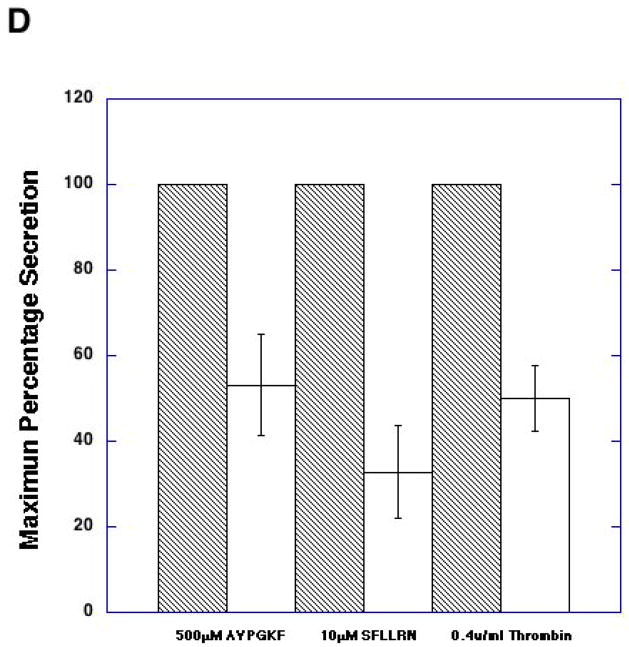
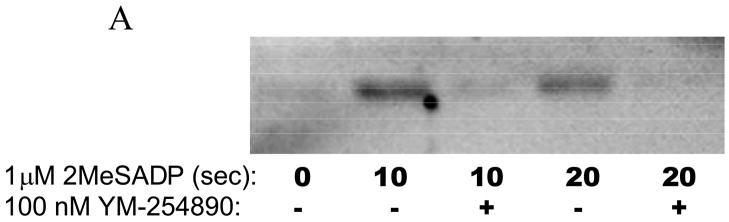
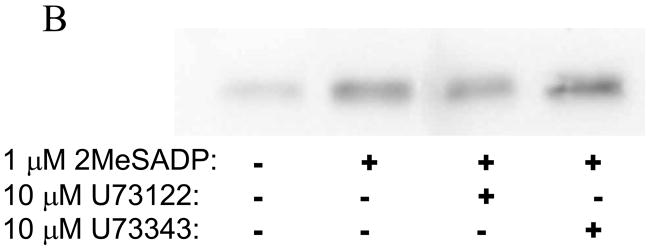
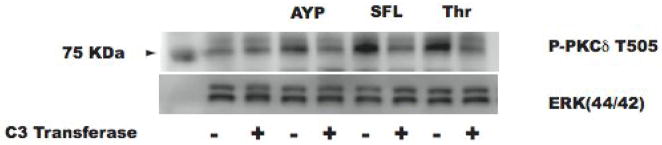
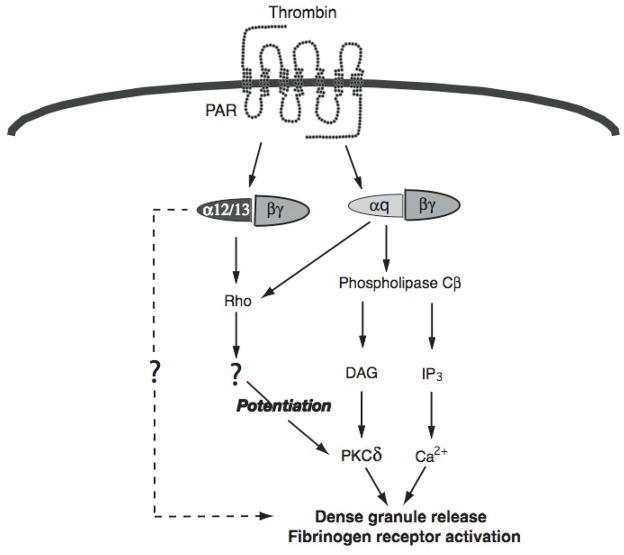
Platelet secretion is an important physiological event in hemostasis. The protease-activated receptors, PAR 1 and PAR 4, and the thromboxane receptor activate the G(12/13) pathways, in addition to the G(q) pathways. Here, we investigated the contribution of G(12/13) pathways to platelet dense granule release. 2MeSADP, which does not activate G(12/13) pathways, does not cause dense granule release in aspirin-treated platelets. However, supplementing 2MeSADP with YFLLRNP (60muM), as selective activator of G(12/13) pathways, resulted in dense granule release. Similarly, supplementing PLC activation with G(12/13) stimulation also leads to dense granule release. These results demonstrate that supplemental signaling from G(12/13) is required for G(q)-mediated dense granule release and that ADP fails to cause dense granule release because the platelet P2Y receptors, although activate PLC, do not activate G(12/13) pathways. When RhoA, downstream signaling molecule in G(12/13) pathways, is blocked, PAR-mediated dense granule release is inhibited. Furthermore, ADP activated RhoA downstream of G(q) and upstream of PLC. Finally, RhoA regulated PKCdelta T505 phosphorylation, suggesting that RhoA pathways contribute to platelet secretion through PKCdelta activation. We conclude that G(12/13) pathways, through RhoA, regulate dense granule release and fibrinogen receptor activation in platelets.
Figures

Similar articles
-
Regulation of protease-activated receptor (PAR) 1 and PAR4 signaling in human platelets by compartmentalized cyclic nucleotide actions.J Pharmacol Exp Ther. 2007 Aug;322(2):778-88. doi: 10.1124/jpet.107.121830. Epub 2007 May 24. J Pharmacol Exp Ther. 2007. PMID: 17525299
-
Protease-activated receptor-induced Akt activation--regulation and possible function.J Thromb Haemost. 2007 Dec;5(12):2484-93. doi: 10.1111/j.1538-7836.2007.02769.x. Epub 2007 Sep 19. J Thromb Haemost. 2007. PMID: 17883592
-
Protease-activated receptors 1 and 4 do not stimulate G(i) signaling pathways in the absence of secreted ADP and cause human platelet aggregation independently of G(i) signaling.Blood. 2002 May 15;99(10):3629-36. doi: 10.1182/blood.v99.10.3629. Blood. 2002. PMID: 11986217
-
Molecular mechanism and functional implications of thrombin-mediated tyrosine phosphorylation of PKCdelta in platelets.Blood. 2005 Jul 15;106(2):550-7. doi: 10.1182/blood-2004-12-4866. Epub 2005 Apr 5. Blood. 2005. PMID: 15811957 Free PMC article.
-
G-protein-coupled receptors as signaling targets for antiplatelet therapy.Arterioscler Thromb Vasc Biol. 2009 Apr;29(4):449-57. doi: 10.1161/ATVBAHA.108.176388. Epub 2008 Nov 20. Arterioscler Thromb Vasc Biol. 2009. PMID: 19023091 Review.
Cited by
-
Neonatal Platelets: Lower G12/13 Expression Contributes to Reduced Secretion of Dense Granules.Cells. 2022 Aug 17;11(16):2563. doi: 10.3390/cells11162563. Cells. 2022. PMID: 36010639 Free PMC article.
-
Negative regulation of Gq-mediated pathways in platelets by G(12/13) pathways through Fyn kinase.J Biol Chem. 2011 Jul 8;286(27):24170-9. doi: 10.1074/jbc.M110.212274. Epub 2011 May 18. J Biol Chem. 2011. PMID: 21592972 Free PMC article.
-
Comprehensive characterization of platelet function in dogs with hyperadrenocorticism.Haematologica. 2025 Aug 1;110(8):1834-1844. doi: 10.3324/haematol.2024.286496. Epub 2025 Feb 6. Haematologica. 2025. PMID: 39911130 Free PMC article.
-
The functional role of protease-activated receptors on contractile responses by activation of Ca2+ sensitization pathways in simian colonic muscles.Am J Physiol Gastrointest Liver Physiol. 2018 Dec 1;315(6):G921-G931. doi: 10.1152/ajpgi.00255.2018. Epub 2018 Sep 27. Am J Physiol Gastrointest Liver Physiol. 2018. PMID: 30260688 Free PMC article.
-
Short-Term E-Cigarette Exposure Increases the Risk of Thrombogenesis and Enhances Platelet Function in Mice.J Am Heart Assoc. 2018 Jul 18;7(15):e009264. doi: 10.1161/JAHA.118.009264. J Am Heart Assoc. 2018. PMID: 30021806 Free PMC article.
References
-
- Packham MA. Role of platelets in thrombosis and hemostasis. Canadian journal of physiology and pharmacology. 1994;72:278–84. - PubMed
-
- Shattil SJ, Kashiwagi H, Pampori N. Integrin signaling: the platelet paradigm. Blood. 1998;91:2645–57. - PubMed
-
- Hourani SM, Cusack NJ. Pharmacological receptors on blood platelets. Pharmacological reviews. 1991;43:243–98. - PubMed
-
- Mills DC. ADP receptors on platelets. Thrombosis and haemostasis. 1996;76:835–56. - PubMed
-
- Molino M, Bainton DF, Hoxie JA, Coughlin SR, Brass LF. Thrombin receptors on human platelets. Initial localization and subsequent redistribution during platelet activation. The Journal of biological chemistry. 1997;272:6011–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
