

Oxidative stress alters syndecan-1 distribution in lungs with pulmonary fibrosis
- PMID: 19073610
- PMCID: PMC2635035
- DOI: 10.1074/jbc.M807001200
Oxidative stress alters syndecan-1 distribution in lungs with pulmonary fibrosis
Abstract
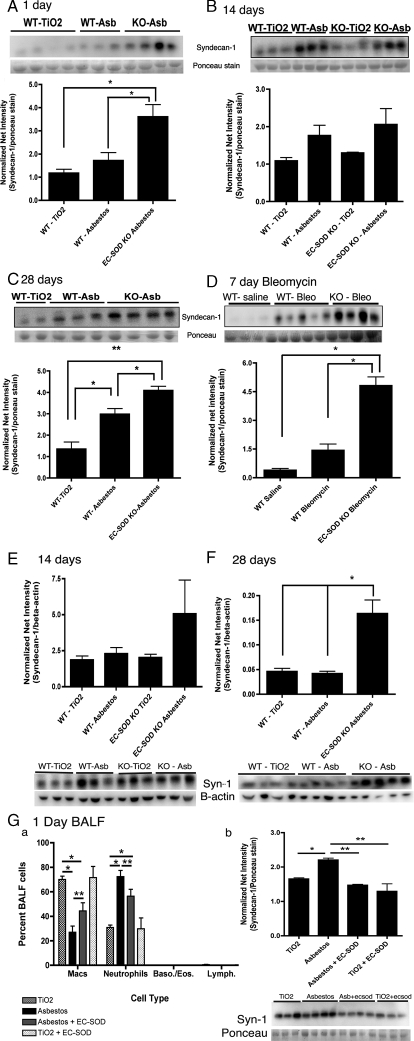
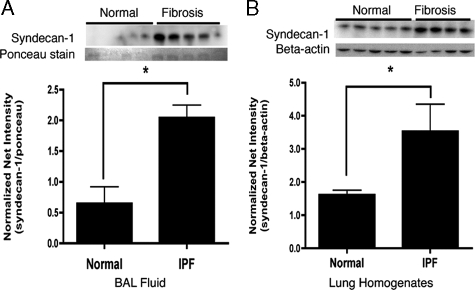
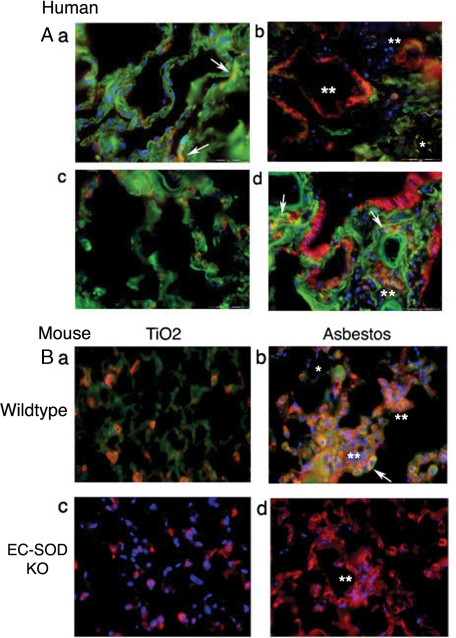
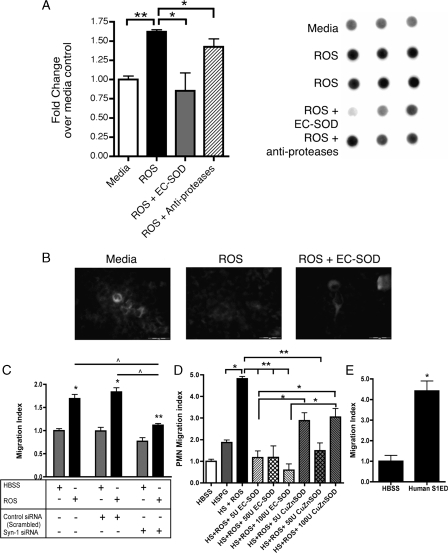
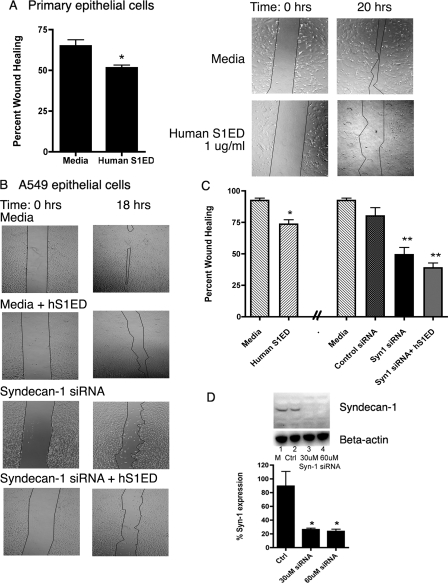
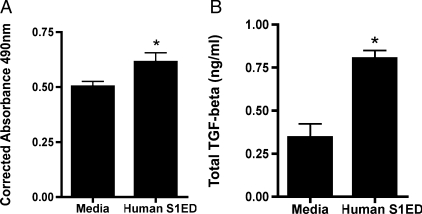
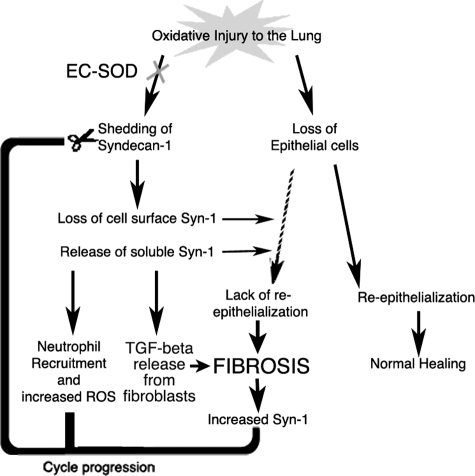
Idiopathic pulmonary fibrosis (IPF) is an interstitial lung disease characterized by severe, progressive fibrosis. Roles for inflammation and oxidative stress have recently been demonstrated, but despite advances in understanding the pathogenesis, there are still no effective therapies for IPF. This study investigates how extracellular superoxide dismutase (EC-SOD), a syndecan-binding antioxidant enzyme, inhibits inflammation and lung fibrosis. We hypothesize that EC-SOD protects the lung from oxidant damage by preventing syndecan fragmentation/shedding. Wild-type or EC-SOD-null mice were exposed to an intratracheal instillation of asbestos or bleomycin. Western blot was used to detect syndecans in the bronchoalveolar lavage fluid and lung. Human lung samples (normal and IPF) were also analyzed. Immunohistochemistry for syndecan-1 and EC-SOD was performed on human and mouse lungs. In vitro, alveolar epithelial cells were exposed to oxidative stress and EC-SOD. Cell supernatants were analyzed for shed syndecan-1 by Western blot. Syndecan-1 ectodomain was assessed in wound healing and neutrophil chemotaxis. Increases in human syndecan-1 are detected in lung homogenates and lavage fluid of IPF lungs. Syndecan-1 is also significantly elevated in the lavage fluid of EC-SOD-null mice after asbestos and bleomycin exposure. On IHC, syndecan-1 staining increases within fibrotic areas of human and mouse lungs. In vitro, EC-SOD inhibits oxidant-induced loss of syndecan-1 from A549 cells. Shed and exogenous syndecan-1 ectodomain induce neutrophil chemotaxis, inhibit alveolar epithelial wound healing, and promote fibrogenesis. Oxidative shedding of syndecan-1 is an underlying cause of neutrophil chemotaxis and aberrant wound healing that may contribute to pulmonary fibrosis.
Figures
References
-
- Dempsey, O. J. (2006) Respir. Med. 100 1871–1885 - PubMed
-
- Flaherty, K. R., Travis, W. D., Colby, T. V., Toews, G. B., Kazerooni, E. A., Gross, B. H., Jain, A., Strawderman, R. L., Flint, A., Lynch, J. P., and Martinez, F. J. (2001) Am. J. Respir. Crit. Care Med. 164 1722–1727 - PubMed
-
- Gross, T. J., and Hunninghake, G. W. (2001) N. Engl. J. Med. 345 517–525 - PubMed
-
- Walter, N., Collard, H. R., and King, T. E., Jr. (2006) Proc. Am. Thorac. Soc. 3 330–338 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
