

Synthesis of CDP-activated ribitol for teichoic acid precursors in Streptococcus pneumoniae
- PMID: 19074383
- PMCID: PMC2631998
- DOI: 10.1128/JB.01120-08
Synthesis of CDP-activated ribitol for teichoic acid precursors in Streptococcus pneumoniae
Abstract
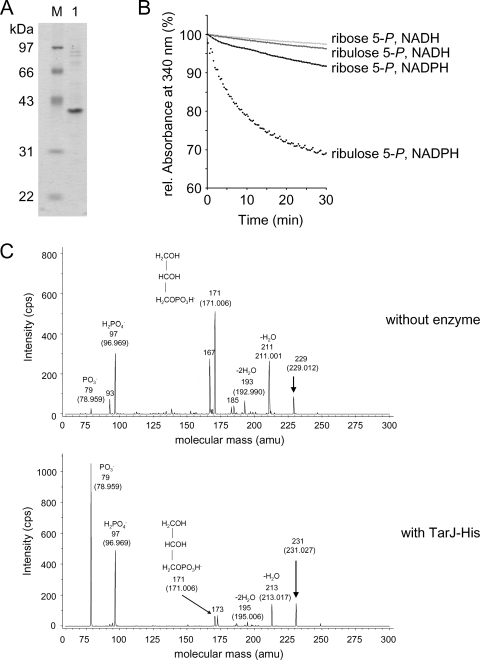
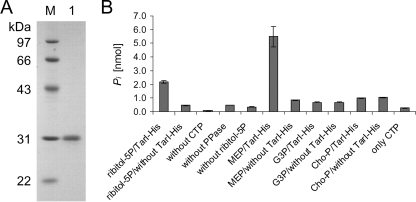
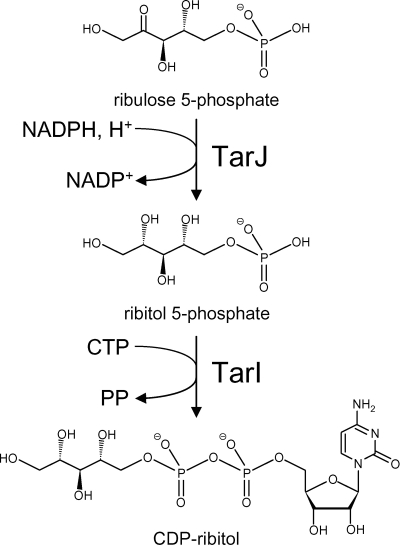
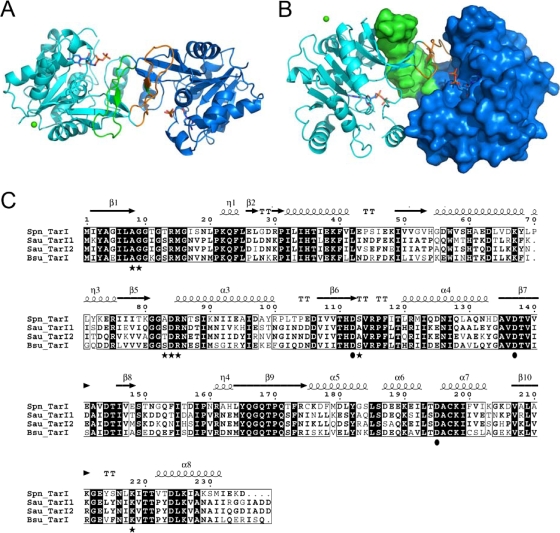
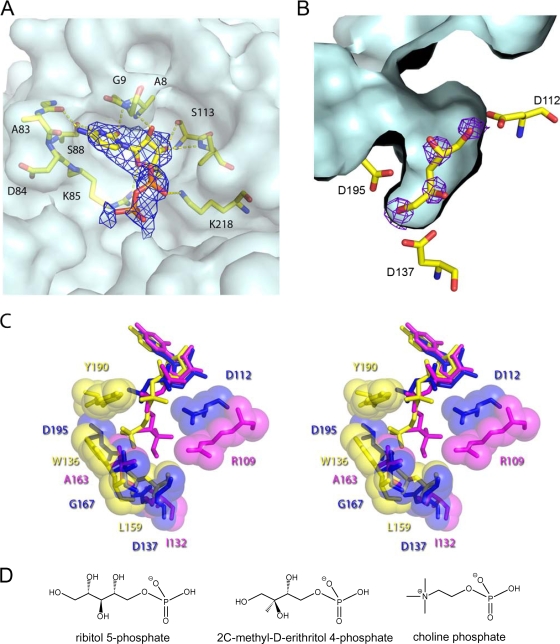
Streptococcus pneumoniae has unusually complex cell wall teichoic acid and lipoteichoic acid, both of which contain a ribitol phosphate moiety. The lic region of the pneumococcal genome contains genes for the uptake and activation of choline, the attachment of phosphorylcholine to teichoic acid precursors, and the transport of these precursors across the cytoplasmic membrane. The role of two other, so far uncharacterized, genes, spr1148 and spr1149, in the lic region was determined. TarJ (spr1148) encodes an NADPH-dependent alcohol dehydrogenase for the synthesis of ribitol 5-phosphate from ribulose 5-phosphate. TarI (spr1149) encodes a cytidylyl transferase for the synthesis of cytidine 5'-diphosphate (CDP)-ribitol from ribitol 5-phosphate and cytidine 5'-triphosphate. We also present the crystal structure of TarI with and without bound CDP, and the structures present a rationale for the substrate specificity of this key enzyme. No transformants were obtained with insertion plasmids designed to interrupt the tarIJ genes, indicating that their function could be essential for cell growth. CDP-activated ribitol is a precursor for the synthesis of pneumococcal teichoic acids and some of the capsular polysaccharides. Thus, all eight genes in the lic region have a role in teichoic acid synthesis.
Figures
References
-
- Badger, J., J. M. Sauder, J. M. Adams, S. Antonysamy, K. Bain, M. G. Bergseid, S. G. Buchanan, M. D. Buchanan, Y. Batiyenko, J. A. Christopher, S. Emtage, A. Eroshkina, I. Feil, E. B. Furlong, K. S. Gajiwala, X. Gao, D. He, J. Hendle, A. Huber, K. Hoda, P. Kearins, C. Kissinger, B. Laubert, H. A. Lewis, J. Lin, K. Loomis, D. Lorimer, G. Louie, M. Maletic, C. D. Marsh, I. Miller, J. Molinari, H. J. Muller-Dieckmann, J. M. Newman, B. W. Noland, B. Pagarigan, F. Park, T. S. Peat, K. W. Post, S. Radojicic, A. Ramos, R. Romero, M. E. Rutter, W. E. Sanderson, K. D. Schwinn, J. Tresser, J. Winhoven, T. A. Wright, L. Wu, J. Xu, and T. J. Harris. 2005. Structural analysis of a set of proteins resulting from a bacterial genomics project. Proteins 60787-796. - PubMed
-
- Baykov, A. A., O. A. Evtushenko, and S. M. Avaeva. 1988. A malachite green procedure for orthophosphate determination and its use in alkaline phosphatase-based enzyme immunoassay. Anal. Biochem. 171266-270. - PubMed
-
- Bentley, S. D., D. M. Aanensen, A. Mavroidi, D. Saunders, E. Rabbinowitsch, M. Collins, K. Donohoe, D. Harris, L. Murphy, M. A. Quail, G. Samuel, I. C. Skovsted, M. S. Kaltoft, B. Barrell, P. R. Reeves, J. Parkhill, and B. G. Spratt. 2006. Genetic analysis of the capsular biosynthetic locus from all 90 pneumococcal serotypes. PLoS Genet. 2e31. - PMC - PubMed
-
- Bernal, C., C. Palacin, A. Boronat, and S. Imperial. 2005. A colorimetric assay for the determination of 4-diphosphocytidyl-2-C-methyl-d-erythritol 4-phosphate synthase activity. Anal. Biochem. 33755-61. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
