

Leucine-responsive regulatory protein (Lrp) acts as a virulence repressor in Salmonella enterica serovar Typhimurium
- PMID: 19074398
- PMCID: PMC2631999
- DOI: 10.1128/JB.01142-08
Leucine-responsive regulatory protein (Lrp) acts as a virulence repressor in Salmonella enterica serovar Typhimurium
Abstract
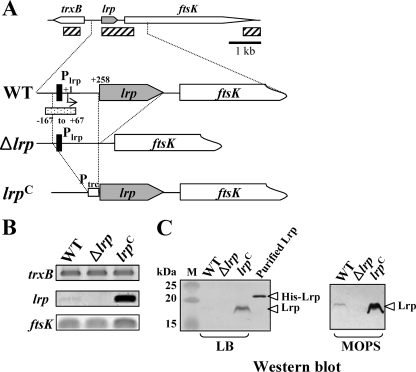
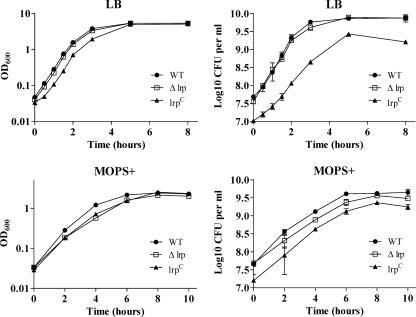
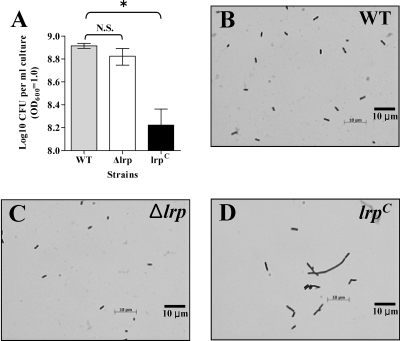
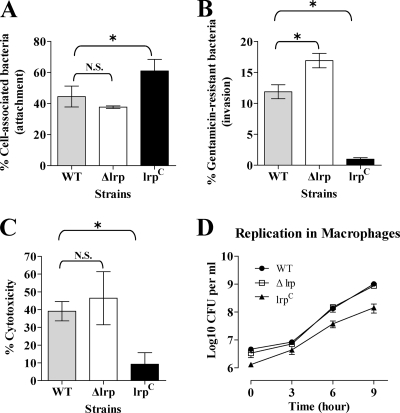
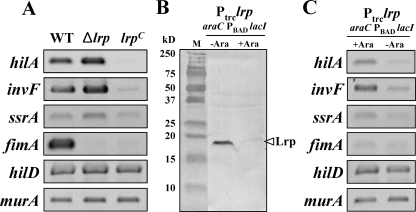
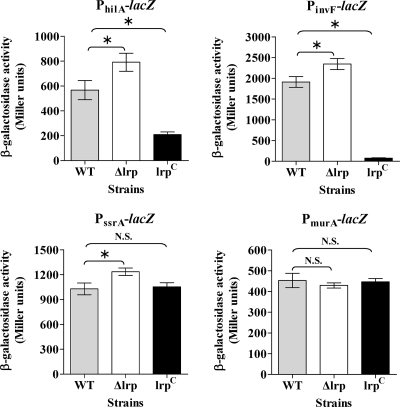
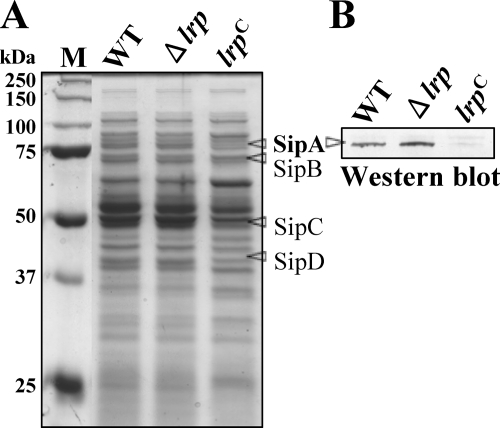
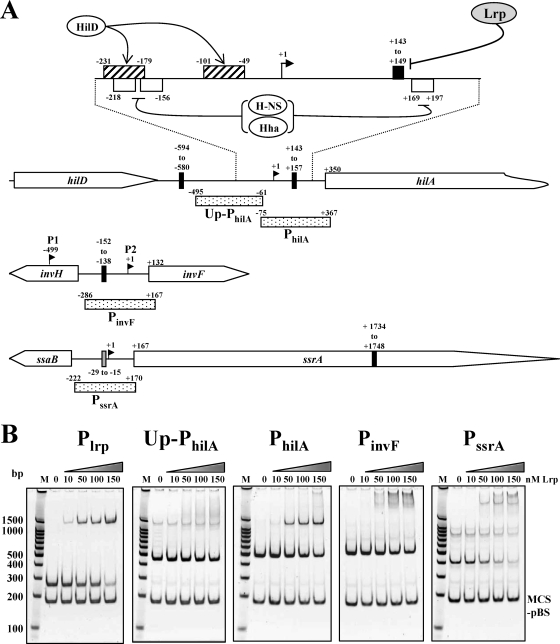
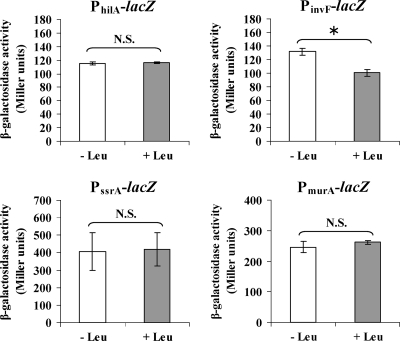
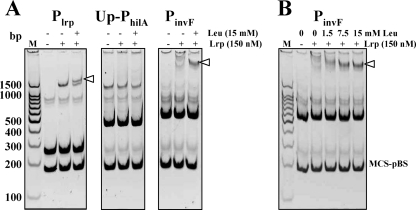
Leucine-responsive regulatory protein (Lrp) is a global gene regulator that influences expression of a large number of genes including virulence-related genes in Escherichia coli and Salmonella. No systematic studies examining the regulation of virulence genes by Lrp have been reported in Salmonella. We report here that constitutive expression of Lrp [lrp(Con)] dramatically attenuates Salmonella virulence while an lrp deletion (Deltalrp) mutation enhances virulence. The lrp(Con) mutant caused pleiotropic effects that include defects in invasion, cytotoxicity, and colonization, whereas the Deltalrp mutant was more proficient at these activities than the wild-type strain. We present evidence that Lrp represses transcription of key virulence regulator genes--hilA, invF, and ssrA--in Salmonella pathogenicity island 1 (SPI-1) and 2 (SPI-2), by binding directly to their promoter regions, P(hilA), P(invF), and P(ssrA). In addition, Western blot analysis showed that the expression of the SPI-1 effector SipA was reduced in the lrp(Con) mutant and enhanced in the Deltalrp mutant. Computational analysis revealed putative Lrp-binding consensus DNA motifs located in P(hilA), P(invF), and P(ssrA). These results suggest that Lrp binds to the consensus motifs and modulates expression of the linked genes. The presence of leucine enhanced Lrp binding to P(invF) in vitro and the addition of leucine to growth medium decreased the level of invF transcription. However, leucine had no effect on expression of hilA and ssrA or on cellular levels of Lrp. In addition, Lrp appears to be an antivirulence gene, since the deletion mutant showed enhanced cell invasion, cytotoxicity, and hypervirulence in BALB/c mice.
Figures
References
-
- Akbar, S., L. M. Schechter, C. P. Lostroh, and C. A. Lee. 2003. AraC/XylS family members, HilD and HilC, directly activate virulence gene expression independently of HilA in Salmonella typhimurium. Mol. Microbiol. 47715-728. - PubMed
-
- Altier, C., M. Suyemoto, A. I. Ruiz, K. D. Burnham, and R. Maurer. 2000. Characterization of two novel regulatory genes affecting Salmonella invasion gene expression. Mol. Microbiol. 35635-646. - PubMed
-
- Arricau, N., D. Hermant, H. Waxin, C. Ecobichon, P. S. Duffey, and M. Y. Popoff. 1998. The RcsB-RcsC regulatory system of Salmonella typhi differentially modulates the expression of invasion proteins, flagellin and Vi antigen in response to osmolarity. Mol. Microbiol. 29835-850. - PubMed
-
- Baek, C.-H., and K.-S. Kim. 2003. lacZ- and aph-based reporter vectors for in vivo expression technology. J. Microbiol. Biotechnol. 13872-880.
-
- Baek, C.-H., K.-E. Lee, D.-K. Park, S.-H. Choi, and K.-S. Kim. 2007. Genetic analysis of spontaneous lactose-utilizing mutants from Vibrio vulnificus. J. Microbiol. Biotechnol. 172046-2055. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
