

Species-specific inhibition of APOBEC3C by the prototype foamy virus protein bet
- PMID: 19074429
- PMCID: PMC2645832
- DOI: 10.1074/jbc.M808853200
Species-specific inhibition of APOBEC3C by the prototype foamy virus protein bet
Abstract
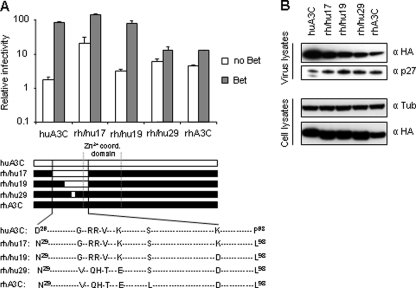
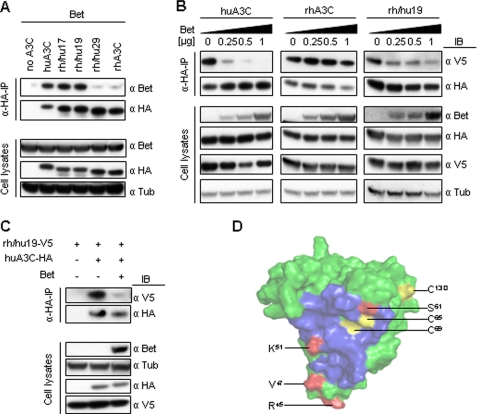
The APOBEC3 cytidine deaminases are part of the intrinsic defense of cells against retroviruses. Lentiviruses and spumaviruses have evolved essential accessory proteins, Vif and Bet, respectively, which counteract the APOBEC3 proteins. We show here that Bet of the Prototype foamy virus inhibits the antiviral APOBEC3C activity by a mechanism distinct to Vif: Bet forms a complex with APOBEC3C without inducing its degradation. Bet abolished APOBEC3C dimerization as shown by coimmunoprecipitation and cross-linking experiments. These findings implicate a physical interaction between Bet and the APOBEC3C. Subsequently, we identified the Bet interaction domain in human APOBEC3C in the predicted APOBEC3C dimerization site. Taken together, these data support the hypothesis that Bet inhibits incorporation of APOBEC3Cs into retroviral particles. Bet likely achieves this by trapping APOBEC3C protein in complexes rendering them unavailable for newly generated viruses due to direct immobilization.
Figures
Similar articles
-
Prototype foamy virus Bet impairs the dimerization and cytosolic solubility of human APOBEC3G.J Virol. 2013 Aug;87(16):9030-40. doi: 10.1128/JVI.03385-12. Epub 2013 Jun 12. J Virol. 2013. PMID: 23760237 Free PMC article.
-
Foamy virus Bet proteins function as novel inhibitors of the APOBEC3 family of innate antiretroviral defense factors.J Virol. 2005 Jul;79(14):8724-31. doi: 10.1128/JVI.79.14.8724-8731.2005. J Virol. 2005. PMID: 15994766 Free PMC article.
-
HIV-2 Vif and foamy virus Bet antagonize APOBEC3B by different mechanisms.Virology. 2021 Feb;554:17-27. doi: 10.1016/j.virol.2020.11.013. Epub 2020 Dec 5. Virology. 2021. PMID: 33333348
-
The APOBEC3 cytidine deaminases: an innate defensive network opposing exogenous retroviruses and endogenous retroelements.Annu Rev Immunol. 2008;26:317-53. doi: 10.1146/annurev.immunol.26.021607.090350. Annu Rev Immunol. 2008. PMID: 18304004 Review.
-
New insights into the role of Vif in HIV-1 replication.AIDS Rev. 2004 Jan-Mar;6(1):34-9. AIDS Rev. 2004. PMID: 15168739 Review.
Cited by
-
Foamy virus budding and release.Viruses. 2013 Apr 10;5(4):1075-98. doi: 10.3390/v5041075. Viruses. 2013. PMID: 23575110 Free PMC article. Review.
-
Phylogeny and History of the Lost SIV from Crab-Eating Macaques: SIVmfa.PLoS One. 2016 Jul 14;11(7):e0159281. doi: 10.1371/journal.pone.0159281. eCollection 2016. PLoS One. 2016. PMID: 27415779 Free PMC article.
-
Basic residues in the foamy virus Gag protein.J Virol. 2011 Apr;85(8):3986-95. doi: 10.1128/JVI.01906-10. Epub 2011 Feb 2. J Virol. 2011. PMID: 21289113 Free PMC article.
-
APOBEC4 Enhances the Replication of HIV-1.PLoS One. 2016 Jun 1;11(6):e0155422. doi: 10.1371/journal.pone.0155422. eCollection 2016. PLoS One. 2016. PMID: 27249646 Free PMC article.
-
Replacement of feline foamy virus bet by feline immunodeficiency virus vif yields replicative virus with novel vaccine candidate potential.Retrovirology. 2018 May 16;15(1):38. doi: 10.1186/s12977-018-0419-0. Retrovirology. 2018. PMID: 29769087 Free PMC article.
References
-
- Bieniasz, P. D. (2004) Nat. Immunol. 5 1109-1115 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
