

doi: 10.1128/EC.00174-08.
Epub 2008 Dec 12.
Functional differentiation of tbf1 orthologues in fission and budding yeasts
Affiliations
- PMID: 19074598
- PMCID: PMC2643609
- DOI: 10.1128/EC.00174-08
Item in Clipboard
Functional differentiation of tbf1 orthologues in fission and budding yeasts
Eukaryot Cell.
2009 Feb.
Abstract
In Saccharomyces cerevisiae, TBF1, an essential gene, influences telomere function but also has other roles in the global regulation of transcription. We have identified a new member of the tbf1 gene family in the mammalian pathogen Pneumocystis carinii. We demonstrate by transspecies complementation that its ectopic expression can provide the essential functions of Schizosaccharomyces pombe tbf1 but that there is no rescue between fission and budding yeast orthologues. Our findings indicate that an essential function of this family of proteins has diverged in the budding and fission yeasts and suggest that effects on telomere length or structure are not the primary cause of inviability in S. pombe tbf1 null strains.
Figures
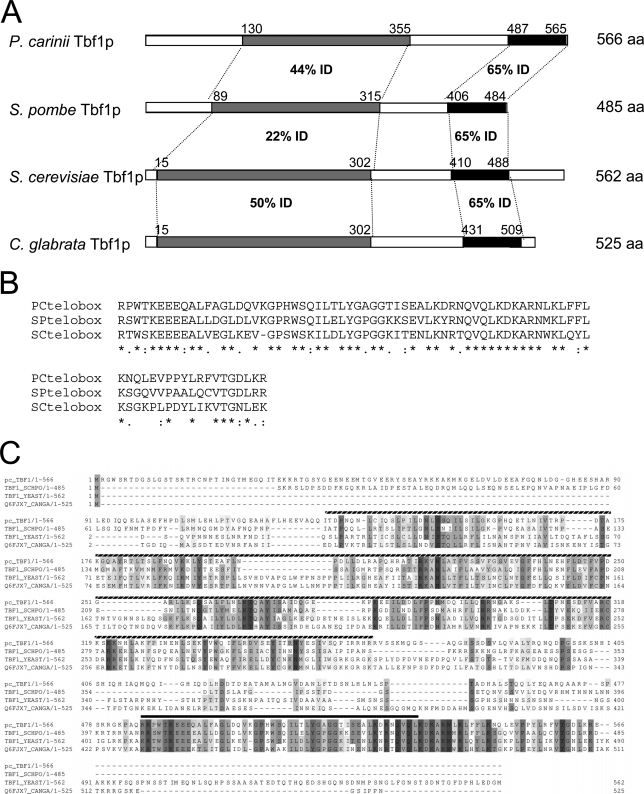
Multiple alignment of P. carinii Tbf1p with other fungal homologues indicates a higher degree of similarity to S. pombe Tbf1p than to S. cerevisiae Tbf1p in the N-terminal domain. (A) A stick diagram illustrating the principal domains of conservation between P. carinii, S. pombe, S. cerevisiae, and C. glabrata Tbf1p. Black boxes indicate the telobox domains: Gray boxes indicate regions corresponding to the less strongly conserved Pfam-B PB010333 domain. (B) Amino acid alignment showing degree of conservation between the telobox domains of P. carinii, S. pombe, and S. cerevisiae Tbf1 proteins. Asterisks indicate positions (65% overall) conserved between all three species. Double dots represent positions containing similar residues. Single dots represent positions containing similar residues in at least two of the sequences. (C) Amino acid alignment showing the conserved domains of Pctbf1, and the S. pombe, S. cerevisiae, and C. glabrata Tbf1 proteins from the Uniprot database. The hatched box indicates the Pfam-B PB010333 domain. The filled box represents the Pfam-A minimal Myb DNA-binding domain. A multiple alignment of Tbf1p sequences from 11 fungal species is shown in Fig. S1 in the supplemental material.
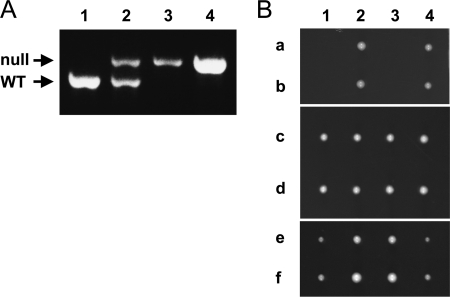
The essential function of S. pombe tbf1 can be provided by expression of P. carinii tbf1 but not S. cerevisiae TBF1. (A) The presence of the tbf1::KanMX4 allele and the absence of the wild-type (WT) Sptbf1 allele in the haploid colonies expressing ectopic Sptbf1 and Pctbf1 was confirmed by PCR using locus-specific primers on genomic DNA from the following strains: 1, wild-type 972 h− haploid strain; 2, heterozygous tbf1/tbf1::KanMX4 diploid; 3, tbf1::KanMX4 haploid expressing pREP41-Sptbf1; 4, tbf1::KanMX4 haploid expressing pREP41-Pctbf1. (B) The lethal phenotype of the S. cerevisiae TBF1::KanMX4 strain is rescued by heterologous expression of S. cerevisiae TBF1 or C. glabrata TBF1 but not by that of P. carinii tbf1. S. cerevisiae diploid cells heterozygous for the TBF1::KanMX4 null allele were transformed with plasmids p416ScTBF1pro-PcTBF1 (rows a and b), p416ScTBF1pro-ScTBF1 (rows c and d), or p416ScTBF1pro-CgTBF1 (rows e and f), and allowed to sporulate. Tetrads from each transformant were separated (columns 1 to 4), grown on rich medium, and then replica plated to verify that viable colonies were dependent on the plasmid.
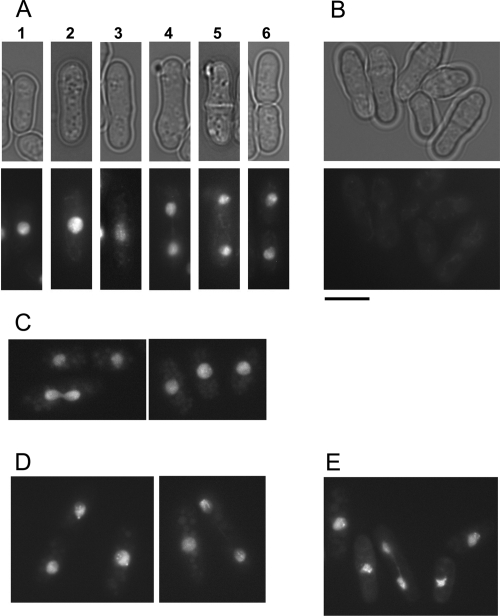
GFP-SpTbf1 GFP-PcTbf1, and GFP-ScTbf1 have similar localization patterns. (A and B) S. pombe leu1::nmt1-41-GFP-Sptbf1(ura4+) cells were grown in the presence (B) or absence (A) of thiamine. At early times after induction (18 h; four generations) GFP-SpTbf1p is observed in the nucleus at all stages of the cell cycle, including early interphase (cell 1), late interphase (cell 2), early mitosis (cell 3), anaphase (cells 4 and 5), and at cytokinesis/G1 (cell 6). Note that in the absence of induction, though the amount of protein produced is sufficient to rescue the tbf1 null allele, the fluorescence of the GFP-tagged protein falls below the threshold of detection. (C) tbf1::KanMX4 leu1::nmt1-41-GFP-Sptbf1(ura4+) cells were grown for 18 h at 25°C in the absence of thiamine. (D) S. pombe tbf1::KanMX4 leu1::nmt1-41-GFP-Pctbf1(ura4+) cells were grown in the absence of thiamine for 18 h at 25°C. (E) leu1::nmt1-41-GFP-Sctbf1(ura4+) cells were grown in the absence of thiamine for 18 h at 25°C. The scale bar represents 10 μm.
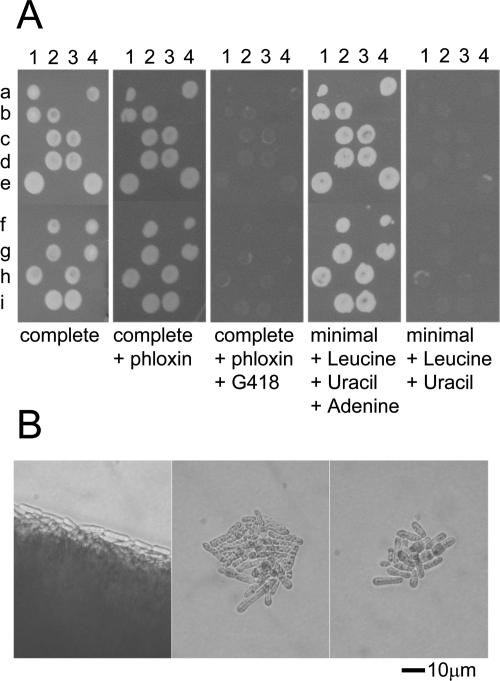
Characterization of the phenotype of Sptbf1 null haploids. An S. pombe diploid heterozygous for the tbf1::KanMX4 null allele was allowed to undergo meiosis, and spores were dissected onto YE medium at 29°C. Colonies were allowed to form and were then replicated to the indicated media. (A) nine tetrads (a to i) are shown; the four spores are at positions 1 to 4. Note that all the surviving colonies are sensitive to G418 and are adenine auxotrophs, indicating that they are haploid, and that cells carrying the tbf1::KanMX4 allele cannot form a visible colony (B). The center and right panels show microcolonies derived from the germinating of tbf1::KanMX4 spores. The null mutant stopped dividing after forming a microcolony of 50 cells or fewer. The cells become elongated compared with cells in a tbf1+ colony, indicative of a cell cycle arrest or delay. For comparison, the left panel shows the edge of a colony of G418-sensitive cells from the same tetrad.
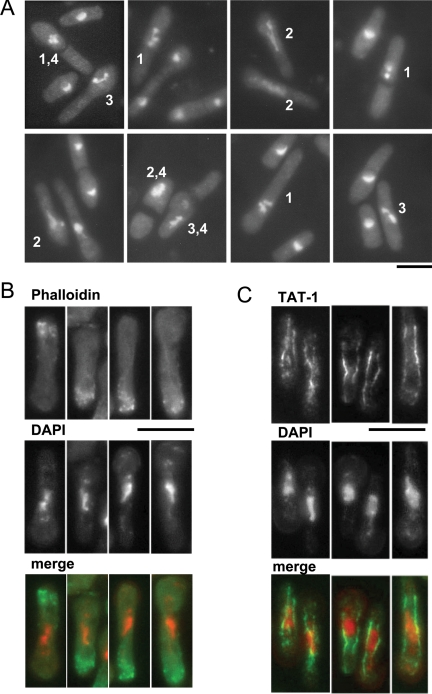
The Sptbf1 null haploids arrest in interphase. Spores prepared from a diploid heterozygous for the tbf1::KanMX4 null allele were grown in complete medium overnight at 19°C, when G418 was added to block further growth and division of tbf1+ cells. At a time corresponding to four generations (see text for details), cells were fixed and the cytoskeleton was analyzed by indirect immunofluorescence. (A) The fixed cells were stained with DAPI. Note the presence of condensed chromosomes (cells marked 1), aberrant nuclear structure (cells marked 2), and problems in chromosome segregation (cells marked 3). Note also the presence of division septa that bisect the cell to generate an anucleate compartment (cells marked 4), indicating that cytokinesis has occurred without nuclear division. (B). The fixed cells were permeabilized and stained with DAPI and rhodamine-conjugated phalloidin. The right panels show tbf1 null cells. Note that F-actin patches are located only at one end of the cell, consistent with a delay early in the cell cycle. (C) The fixed cells were digested, and the TAT-1 antibody was used to analyze the microtubules, as described in Materials and Methods. DAPI is shown in red and microtubules in green. The right panels show the phenotype of elongated tbf1 null cells. Note that all of them have an interphase microtubule array. The scale bars for each panel represent 10 μm.
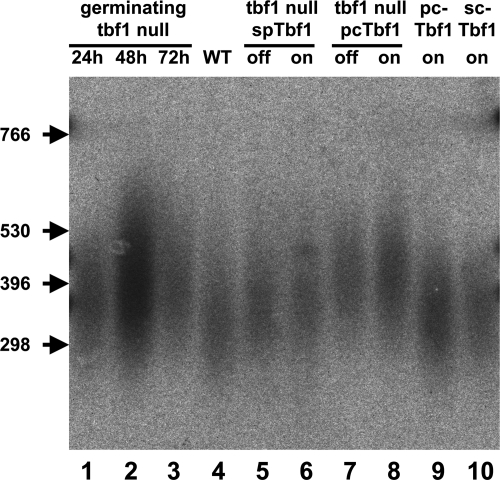
Telomere length in both tbf1 null cells and cells rescued by GFP-SpTbf1 or GFP-PcTbf1 is slightly longer than wild type. Genomic DNA was extracted from the indicated strains and digested overnight with ApaI. The DNA fragments were separated by electrophoresis overnight at 20 V (1 V/cm) through a 1.5% (wt/vol) agarose gel. The filter was probed with a synthetic telomere fragment as described in Materials and Methods. A portion of the filter is shown. The arrowheads indicate the positions of DNA markers visualized by ethidium bromide staining prior to transfer. The lane numbers are referred to in the text. Lanes 1 to 3, DNA from germinating tbf1 null spores at 24, 48, and 72 h after germination. Note that wild-type (WT) cells do not contribute significantly to the culture, as they are unable to divide in the presence of G418, while null spores undergo multiple divisions. Lane 4, wild-type cells. Lanes 5 and 6, DNA from tbf1::KanMX4 leu1::nmt1-41-GFP-Sptbf1(ura4+) cells grown in the absence (on) or presence (off) of thiamine. Lanes 7 and 8, DNA from tbf1::KanMX4 leu1::nmt1-41-GFP-Pctbf1(ura4+) cells grown in the absence (on) or presence (off) of thiamine. Lane 9, DNA from leu1::nmt1-41-GFP-Pctbf1(ura4+) cells grown in the absence (on) of thiamine. Lane 10, DNA from leu1::nmt1-41-GFP-Sctbf1(ura4+) cells grown in the absence (on) of thiamine. Note that lane 2 contains approximately three times as much DNA as the others.
Similar articles
-
Involvement of rhp23, a Schizosaccharomyces pombe homolog of the human HHR23A and Saccharomyces cerevisiae RAD23 nucleotide excision repair genes, in cell cycle control and protein ubiquitination.Nucleic Acids Res. 2002 Jan 15;30(2):581-91. doi: 10.1093/nar/30.2.581. Nucleic Acids Res. 2002. PMID: 11788722 Free PMC article.
-
Structural and functional insights into yeast Tbf1 as an atypical telomeric repeat-binding factor.Structure. 2024 Jul 11;32(7):889-898.e3. doi: 10.1016/j.str.2024.04.002. Epub 2024 Apr 26. Structure. 2024. PMID: 38677290
-
Identification and characterization of an essential telomeric repeat binding factor in fission yeast.J Biol Chem. 2008 Feb 1;283(5):2693-701. doi: 10.1074/jbc.M708784200. Epub 2007 Oct 30. J Biol Chem. 2008. PMID: 17977837
-
Genetic approaches to aging in budding and fission yeasts: new connections and new opportunities.Subcell Biochem. 2012;57:291-314. doi: 10.1007/978-94-007-2561-4_13. Subcell Biochem. 2012. PMID: 22094427 Review.
-
A conserved role of the RSC chromatin remodeler in the establishment of nucleosome-depleted regions.Curr Genet. 2017 May;63(2):187-193. doi: 10.1007/s00294-016-0642-y. Epub 2016 Aug 24. Curr Genet. 2017. PMID: 27558480 Free PMC article. Review.
Cited by
-
Tbf1 and Vid22 promote resection and non-homologous end joining of DNA double-strand break ends.EMBO J. 2013 Jan 23;32(2):275-89. doi: 10.1038/emboj.2012.327. Epub 2012 Dec 7. EMBO J. 2013. PMID: 23222485 Free PMC article.
-
Functional characterization of the Pneumocystis jirovecii potential drug targets dhfs and abz2 involved in folate biosynthesis.Antimicrob Agents Chemother. 2015 May;59(5):2560-6. doi: 10.1128/AAC.05092-14. Epub 2015 Feb 17. Antimicrob Agents Chemother. 2015. PMID: 25691634 Free PMC article.
-
Subtelomere-binding protein Tbf1 and telomere-binding protein Rap1 collaborate to inhibit localization of the Mre11 complex to DNA ends in budding yeast.Mol Biol Cell. 2012 Jan;23(2):347-59. doi: 10.1091/mbc.E11-06-0568. Epub 2011 Nov 30. Mol Biol Cell. 2012. PMID: 22130795 Free PMC article.
-
Evidence for Proinflammatory β-1,6 Glucans in the Pneumocystis carinii Cell Wall.Infect Immun. 2015 Jul;83(7):2816-26. doi: 10.1128/IAI.00196-15. Epub 2015 Apr 27. Infect Immun. 2015. PMID: 25916991 Free PMC article.
-
Tay1 protein, a novel telomere binding factor from Yarrowia lipolytica.J Biol Chem. 2010 Dec 3;285(49):38078-92. doi: 10.1074/jbc.M110.127605. Epub 2010 Oct 5. J Biol Chem. 2010. PMID: 20923774 Free PMC article.
References
-
- Ambrose, H. E., S. P. Keely, E. M. Aliouat, E. Dei-Cas, A. E. Wakefield, R. F. Miller, and J. R. Stringer. 2004. Expression and complexity of the PRT1 multigene family of Pneumocystis carinii. Microbiology 150293-300. - PubMed
-
- Aslett, M., and V. Wood. 2006. Gene Ontology annotation status of the fission yeast genome: preliminary coverage approaches 100%. Yeast 23913-919. - PubMed
-
- Barry, J. D., M. L. Ginger, P. Burton, and R. McCulloch. 2003. Why are parasite contingency genes often associated with telomeres? Int. J. Parasitol. 3329-45. - PubMed
-
- Basi, G., E. Schmid, and K. Maundrell. 1993. TATA box mutations in the Schizosaccharomyces pombe nmt1 promoter affect transcription efficiency but not the transcription start point or thiamine repressibility. Gene 123131-136. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
