

Aurora-A kinase is essential for bipolar spindle formation and early development
- PMID: 19075002
- PMCID: PMC2643803
- DOI: 10.1128/MCB.01062-08
Aurora-A kinase is essential for bipolar spindle formation and early development
Abstract
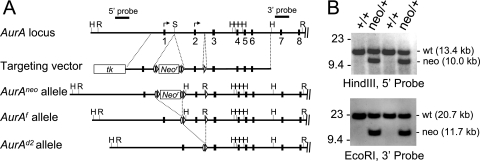
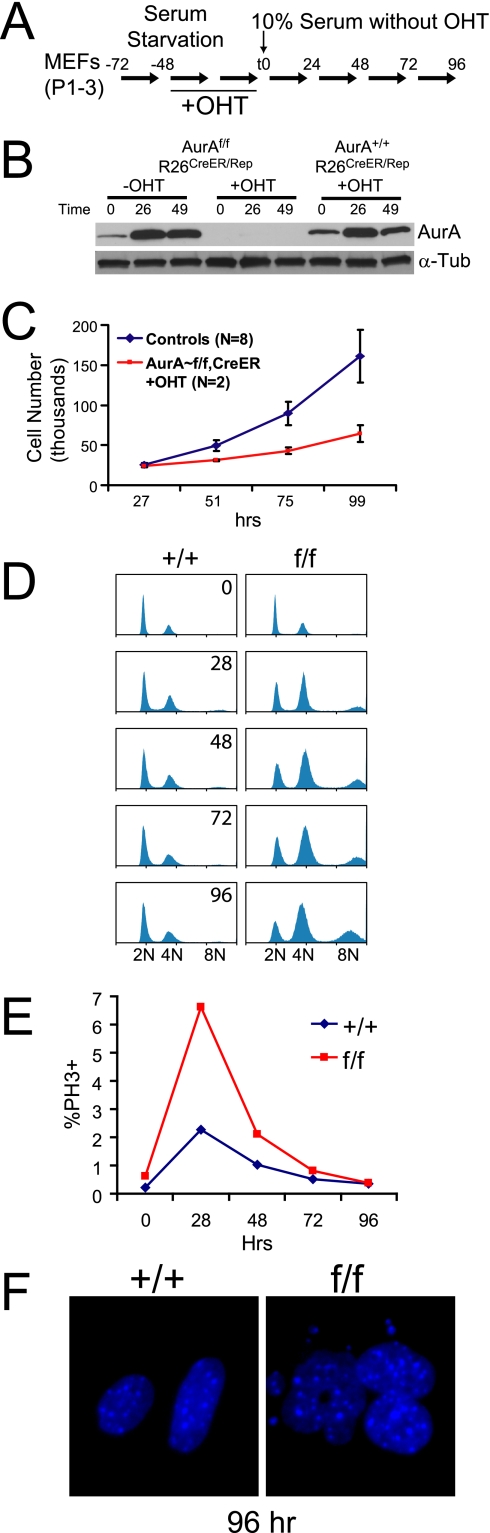
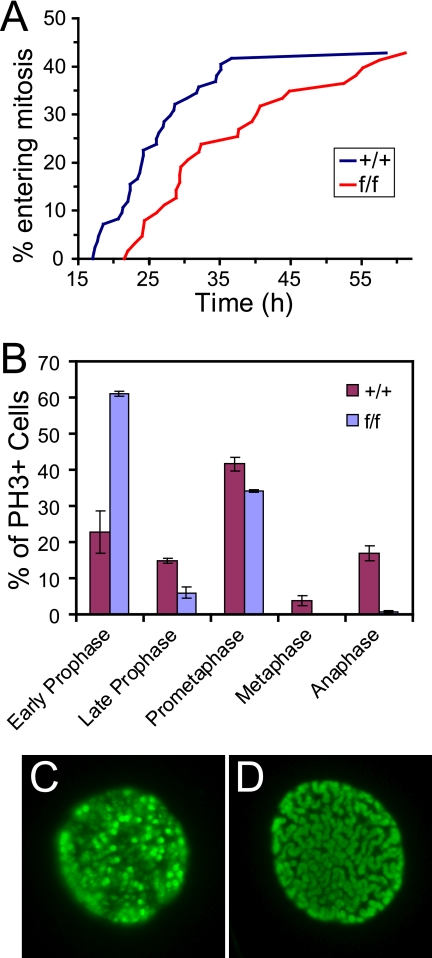
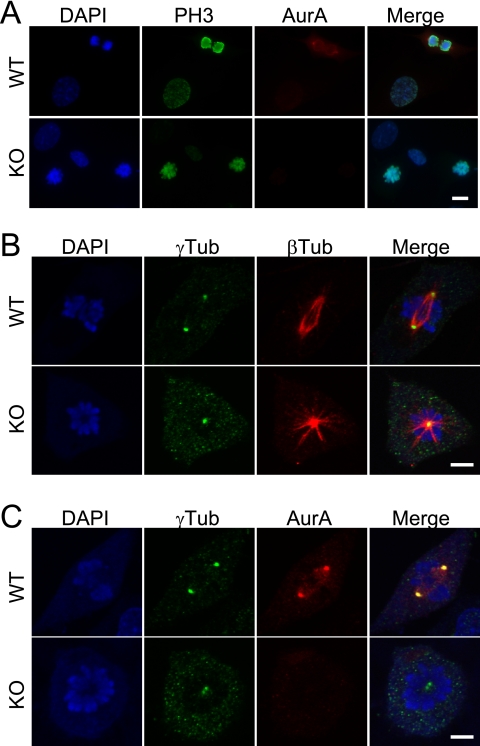
Aurora-A is a conserved kinase implicated in mitotic regulation and carcinogenesis. Aurora-A was previously implicated in mitotic entry and spindle assembly, although contradictory results prevented a clear understanding of the roles of Aurora-A in mammals. We developed a conditional null mutation in the mouse Aurora-A gene to investigate Aurora-A functions in primary cells ex vivo and in vivo. We show here that conditional Aurora-A ablation in cultured embryonic fibroblasts causes impaired mitotic entry and mitotic arrest with a profound defect in bipolar spindle formation. Germ line Aurora-A deficiency causes embryonic death at the blastocyst stage with pronounced cell proliferation failure, mitotic arrest, and monopolar spindle formation. Aurora-A deletion in mid-gestation embryos causes an increase in mitotic and apoptotic cells. These results indicate that murine Aurora-A facilitates, but is not absolutely required for, mitotic entry in murine embryonic fibroblasts and is essential for centrosome separation and bipolar spindle formation in vitro and in vivo. Aurora-A deletion increases apoptosis, suggesting that molecular therapies targeting Aurora-A may be effective in inducing tumor cell apoptosis. Aurora-A conditional mutant mice provide a valuable system for further defining Aurora-A functions and for predicting effects of Aurora-A therapeutic intervention.
Figures
References
-
- Andrews, P. D., E. Knatko, W. J. Moore, and J. R. Swedlow. 2003. Mitotic mechanics: the auroras come into view. Curr. Opin. Cell Biol. 15672-683. - PubMed
-
- Berdnik, D., and J. A. Knoblich. 2002. Drosophila Aurora-A is required for centrosome maturation and actin-dependent asymmetric protein localization during mitosis. Curr. Biol. 12640-647. - PubMed
-
- Bergnes, G., K. Brejc, and L. Belmont. 2005. Mitotic kinesins: prospects for antimitotic drug discovery. Curr. Top. Med. Chem. 5127-145. - PubMed
-
- Bischoff, J. R., L. Anderson, Y. Zhu, K. Mossie, L. Ng, B. Souza, B. Schryver, P. Flanagan, F. Clairvoyant, C. Ginther, C. S. Chan, M. Novotny, D. J. Slamon, and G. D. Plowman. 1998. A homologue of Drosophila aurora kinase is oncogenic and amplified in human colorectal cancers. EMBO J. 173052-3065. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials