

A postnatal switch of CELF and MBNL proteins reprograms alternative splicing in the developing heart
- PMID: 19075228
- PMCID: PMC2629332
- DOI: 10.1073/pnas.0809045105
A postnatal switch of CELF and MBNL proteins reprograms alternative splicing in the developing heart
Abstract
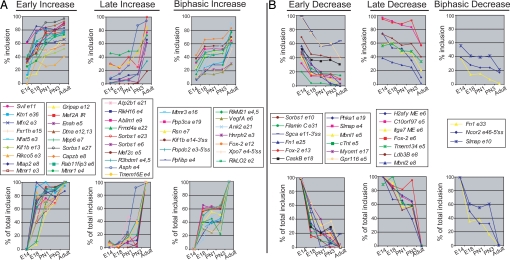
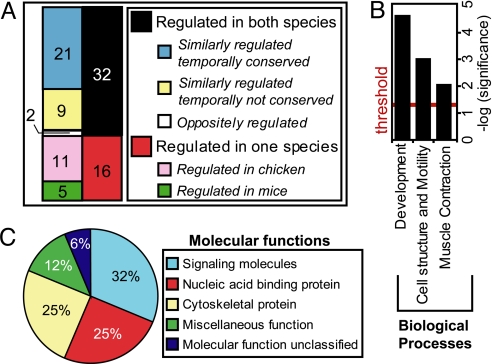
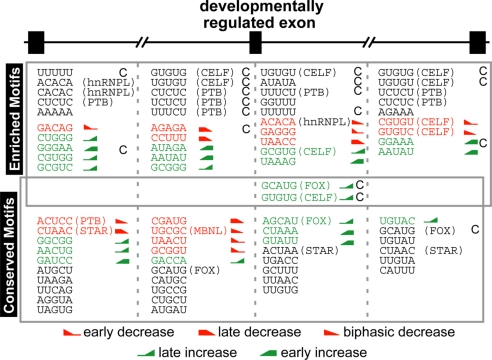
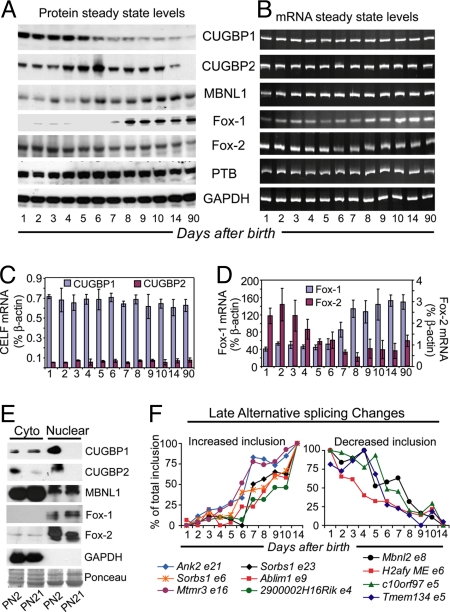
From a large-scale screen using splicing microarrays and RT-PCR, we identified 63 alternative splicing (AS) events that are coordinated in 3 distinct temporal patterns during mouse heart development. More than half of these splicing transitions are evolutionarily conserved between mouse and chicken. Computational analysis of the introns flanking these splicing events identified enriched and conserved motifs including binding sites for CUGBP and ETR-3-like factors (CELF), muscleblind-like (MBNL) and Fox proteins. We show that CELF proteins are down-regulated >10-fold during heart development, and MBNL1 protein is concomitantly up-regulated nearly 4-fold. Using transgenic and knockout mice, we show that reproducing the embryonic expression patterns for CUGBP1 and MBNL1 in adult heart induces the embryonic splicing patterns for more than half of the developmentally regulated AS transitions. These findings indicate that CELF and MBNL proteins are determinative for a large subset of splicing transitions that occur during postnatal heart development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Graveley BR. Alternative splicing: Increasing diversity in the proteomic world. Trends Genet. 2001;17:100–107. - PubMed
-
- Johnson JM, et al. Genome-wide survey of human alternative pre-mRNA splicing with exon junction microarrays. Science. 2003;302:2141–2144. - PubMed
-
- Wang GS, Cooper TA. Splicing in disease: Disruption of the splicing code and the decoding machinery. Nat Rev Genet. 2007;8:749–761. - PubMed
-
- Gabut M, Chaudhry S, Blencowe BJ. SnapShot: The splicing regulatory machinery. Cell. 2008;133:192–e1. - PubMed
-
- Matlin AJ, Clark F, Smith CW. Understanding alternative splicing: Towards a cellular code. Nat Rev Mol Cell Biol. 2005;6:386–398. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
