

TLR9 traffics through the Golgi complex to localize to endolysosomes and respond to CpG DNA
- PMID: 19079358
- PMCID: PMC2753824
- DOI: 10.1038/icb.2008.101
TLR9 traffics through the Golgi complex to localize to endolysosomes and respond to CpG DNA
Abstract
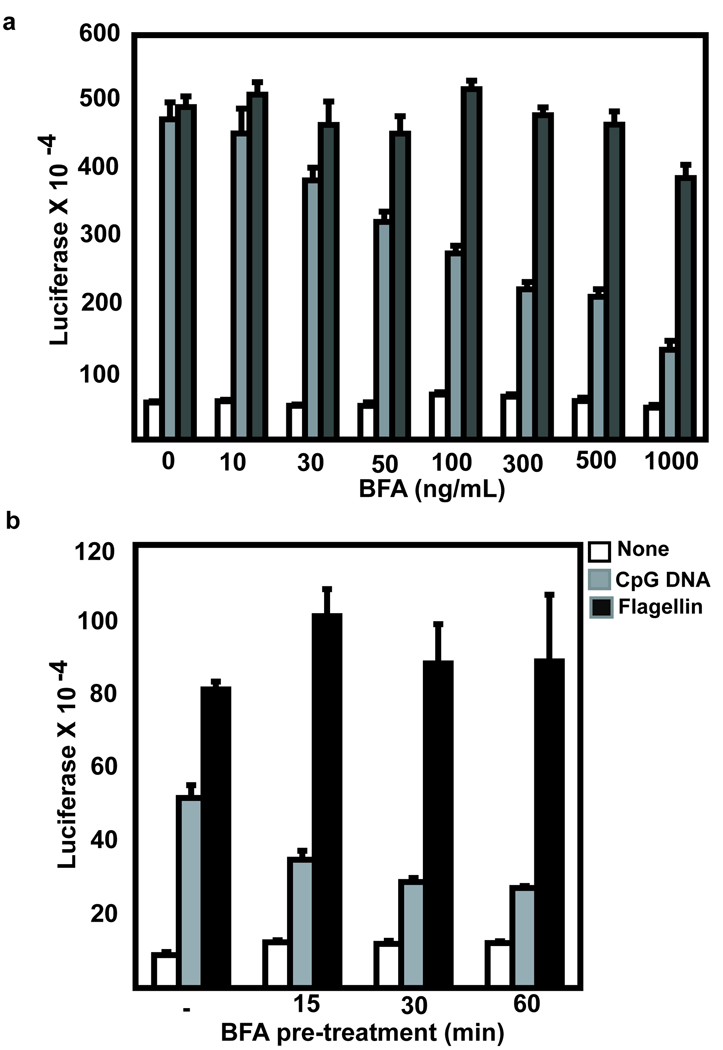
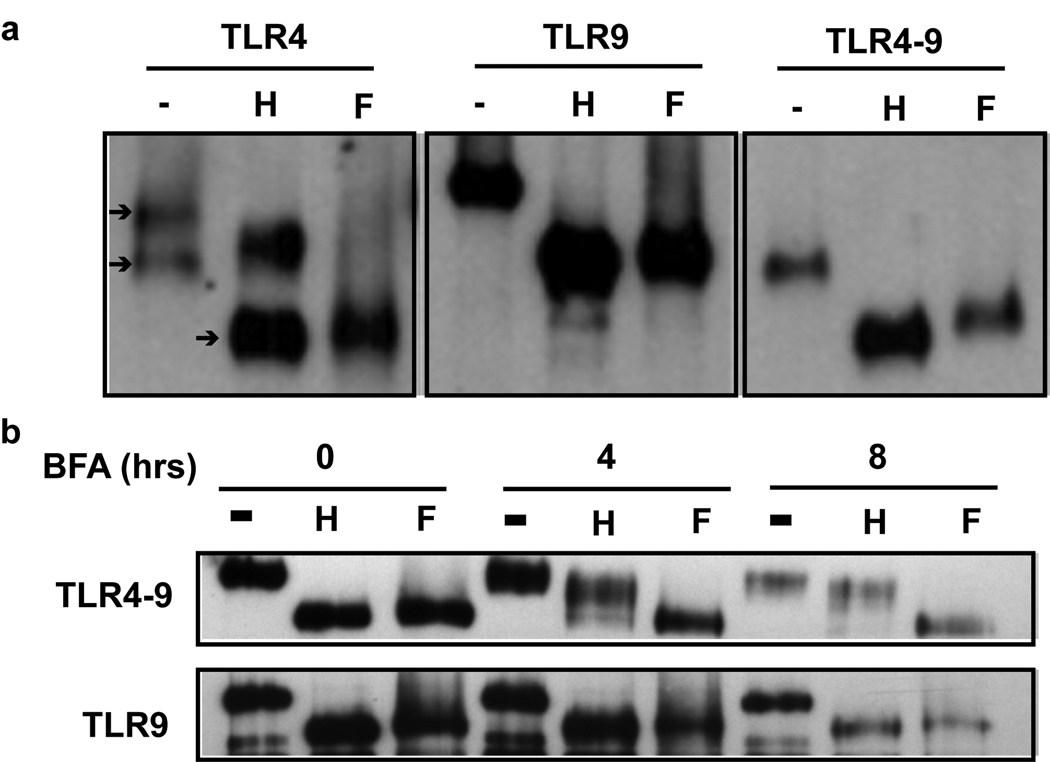
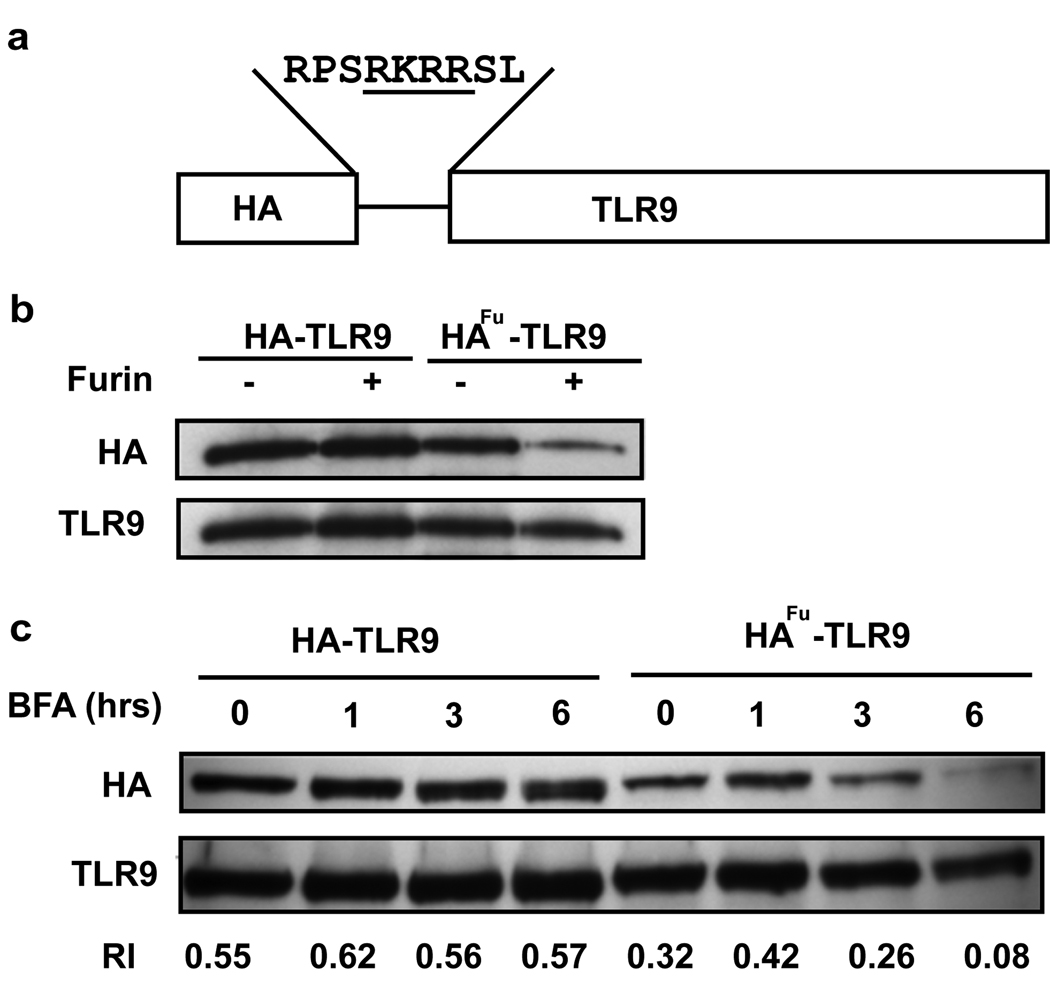
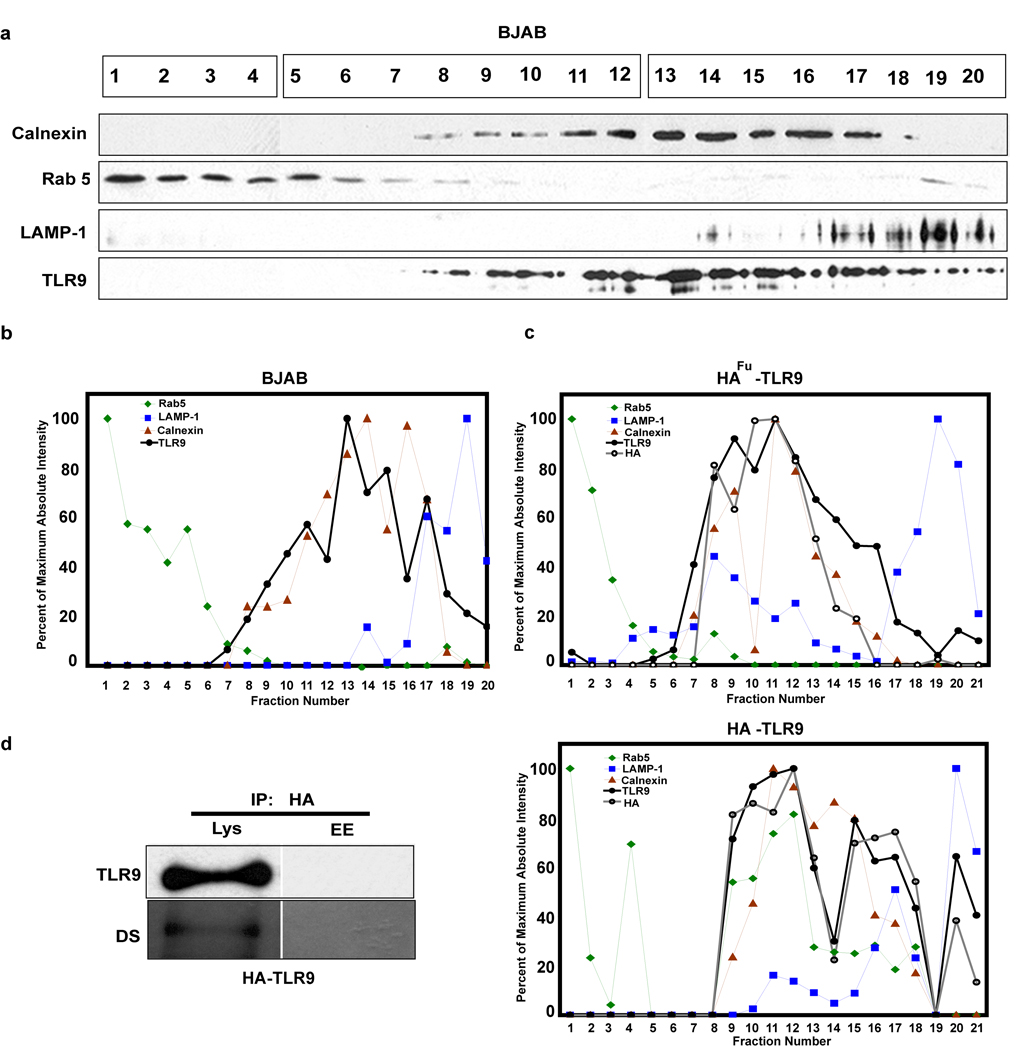
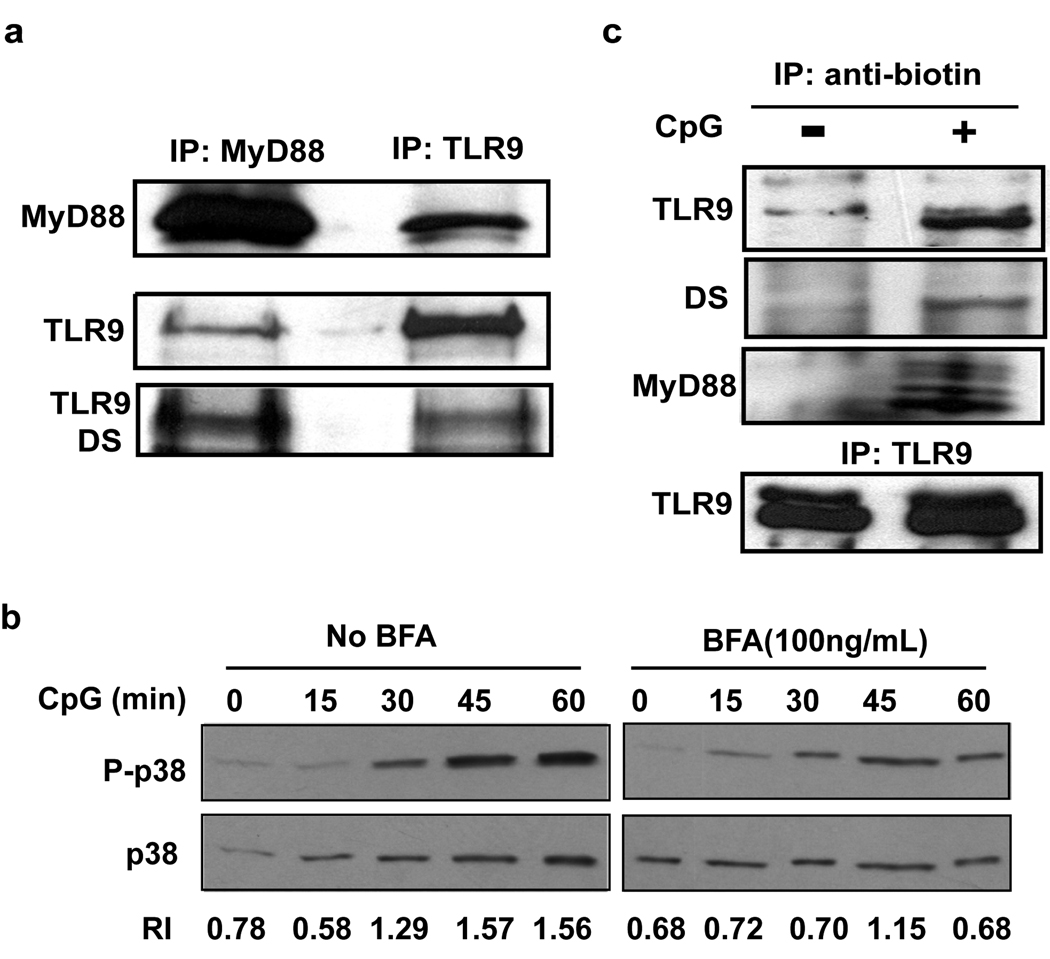
Toll-like receptor 9 (TLR9) promiscuously binds self- and microbial DNA, but only microbial DNA elicits an inflammatory response. How TLR9 discriminates between self- and foreign DNA is unclear, but inappropriate localization of TLR9 permits response to self-DNA, suggesting that TLR9 localization and trafficking are critical components. The molecular mechanisms controlling the movement of TLR9 may provide new insight into the recognition of DNA in normal and in pathological conditions such as autoimmune systemic lupus erythematosus. We have shown earlier that TLR9 is retained in the endoplasmic reticulum (ER) and it moves to endolysosomes to recognize CpG DNA. Other studies have suggested that TLR9 bypasses the Golgi complex to access endolysosomes. Here, we show that TLR9 translocates from ER to endolysosomes through the Golgi complex and that Golgi export is required for optimal TLR9 signaling. In all, 6-13% of TLR9 constitutively exits the ER, moves through the Golgi complex and resides in lysosomal-associated membrane protein-1-positive vesicles. TLR9 bound to CpG DNA had glycan modifications indicative of Golgi processing confirming that TLR9 travels through the Golgi complex to access CpG DNA in endolysosomes. Together, these data support a model where TLR9 uses traditional secretory pathways and does not bypass the Golgi complex.
Figures

References
-
- Takeda K, Takeuchi O, Akira S. Recognition of lipopeptides by Toll-like receptors. J. Endotoxin Res. 2002;8:459–463. - PubMed
-
- Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature. 2001;413:732–738. - PubMed
-
- Heil F, Hemmi H, Hochrein H, Ampenberger F, Kirschning C, Akira S, et al. Species-specific recognition of single-stranded RNA via Toll-like receptor 7 and 8. Science. 2004;303:1526–1529. - PubMed
-
- Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H, et al. A Toll-like receptor recognizes bacterial DNA. Nature. 2000;408:740–745. - PubMed
-
- Takeshita F, Leifer CA, Gursel I, Ishii KJ, Takeshita S, Gursel M, et al. Cutting edge: Role of Toll-like receptor 9 in CpG DNA-induced activation of human cells. J. Immunol. 2001;167:3555–3558. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
