

The repertoire and dynamics of evolutionary adaptations to controlled nutrient-limited environments in yeast
- PMID: 19079573
- PMCID: PMC2586090
- DOI: 10.1371/journal.pgen.1000303
The repertoire and dynamics of evolutionary adaptations to controlled nutrient-limited environments in yeast
Abstract
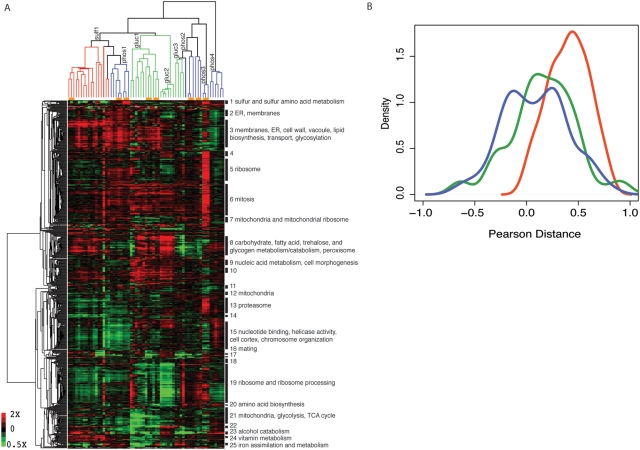
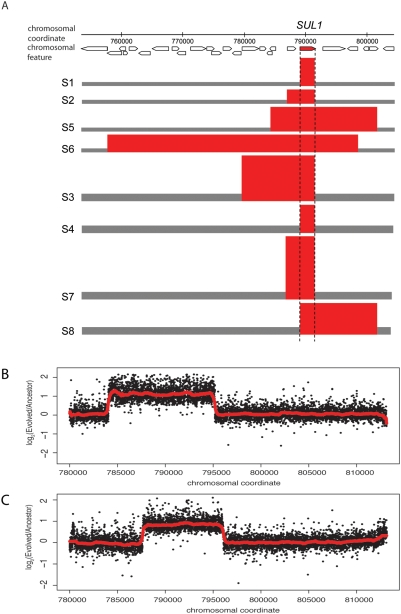
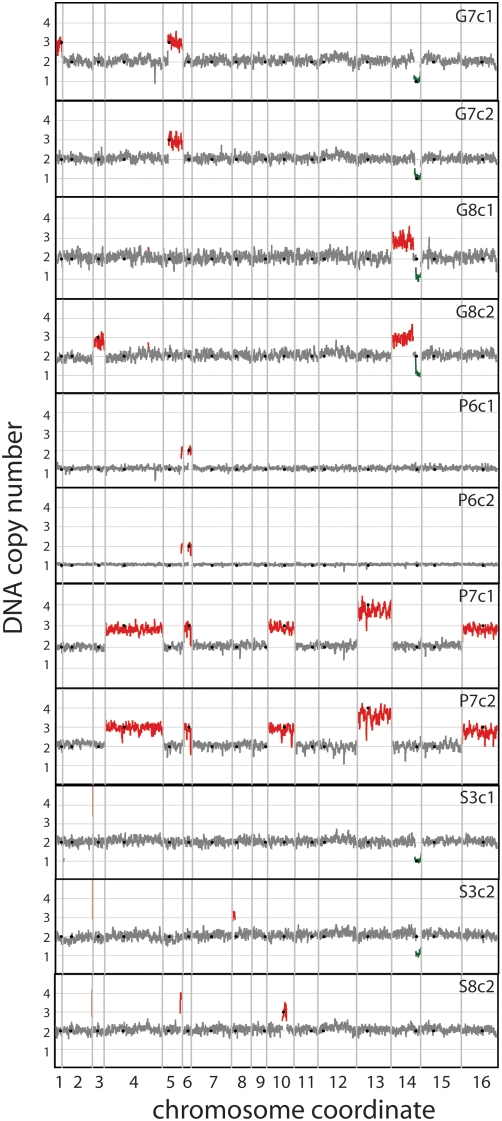
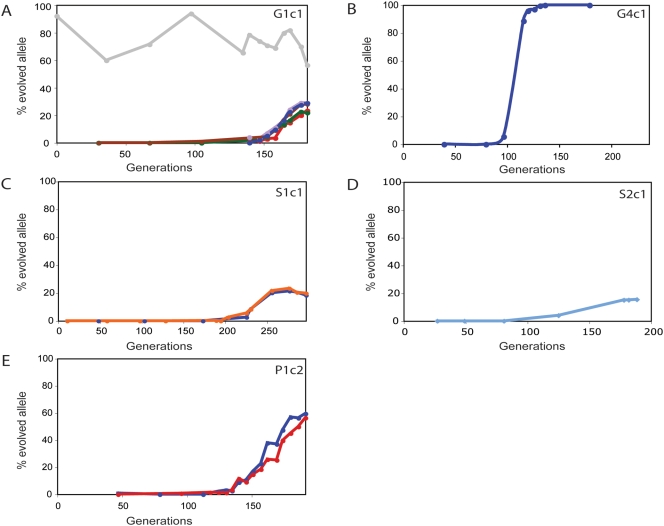
The experimental evolution of laboratory populations of microbes provides an opportunity to observe the evolutionary dynamics of adaptation in real time. Until very recently, however, such studies have been limited by our inability to systematically find mutations in evolved organisms. We overcome this limitation by using a variety of DNA microarray-based techniques to characterize genetic changes -- including point mutations, structural changes, and insertion variation -- that resulted from the experimental adaptation of 24 haploid and diploid cultures of Saccharomyces cerevisiae to growth in either glucose, sulfate, or phosphate-limited chemostats for approximately 200 generations. We identified frequent genomic amplifications and rearrangements as well as novel retrotransposition events associated with adaptation. Global nucleotide variation detection in ten clonal isolates identified 32 point mutations. On the basis of mutation frequencies, we infer that these mutations and the subsequent dynamics of adaptation are determined by the batch phase of growth prior to initiation of the continuous phase in the chemostat. We relate these genotypic changes to phenotypic outcomes, namely global patterns of gene expression, and to increases in fitness by 5-50%. We found that the spectrum of available mutations in glucose- or phosphate-limited environments combined with the batch phase population dynamics early in our experiments allowed several distinct genotypic and phenotypic evolutionary pathways in response to these nutrient limitations. By contrast, sulfate-limited populations were much more constrained in both genotypic and phenotypic outcomes. Thus, the reproducibility of evolution varies with specific selective pressures, reflecting the constraints inherent in the system-level organization of metabolic processes in the cell. We were able to relate some of the observed adaptive mutations (e.g., transporter gene amplifications) to known features of the relevant metabolic pathways, but many of the mutations pointed to genes not previously associated with the relevant physiology. Thus, in addition to answering basic mechanistic questions about evolutionary mechanisms, our work suggests that experimental evolution can also shed light on the function and regulation of individual metabolic pathways.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Bollback JP, Huelsenbeck JP. Clonal interference is alleviated by high mutation rates in large populations. Mol Biol Evol. 2007;24:1397–1406. - PubMed
-
- Herring CD, Raghunathan A, Honisch C, Patel T, Applebee MK, et al. Comparative genome sequencing of Escherichia coli allows observation of bacterial evolution on a laboratory timescale. Nat Genet. 2006;38:1406–1412. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
