

Heterosubtypic neutralizing monoclonal antibodies cross-protective against H5N1 and H1N1 recovered from human IgM+ memory B cells
- PMID: 19079604
- PMCID: PMC2596486
- DOI: 10.1371/journal.pone.0003942
Heterosubtypic neutralizing monoclonal antibodies cross-protective against H5N1 and H1N1 recovered from human IgM+ memory B cells
Abstract
Background: The hemagglutinin (HA) glycoprotein is the principal target of protective humoral immune responses to influenza virus infections but such antibody responses only provide efficient protection against a narrow spectrum of HA antigenic variants within a given virus subtype. Avian influenza viruses such as H5N1 are currently panzootic and pose a pandemic threat. These viruses are antigenically diverse and protective strategies need to cross protect against diverse viral clades. Furthermore, there are 16 different HA subtypes and no certainty the next pandemic will be caused by an H5 subtype, thus it is important to develop prophylactic and therapeutic interventions that provide heterosubtypic protection.
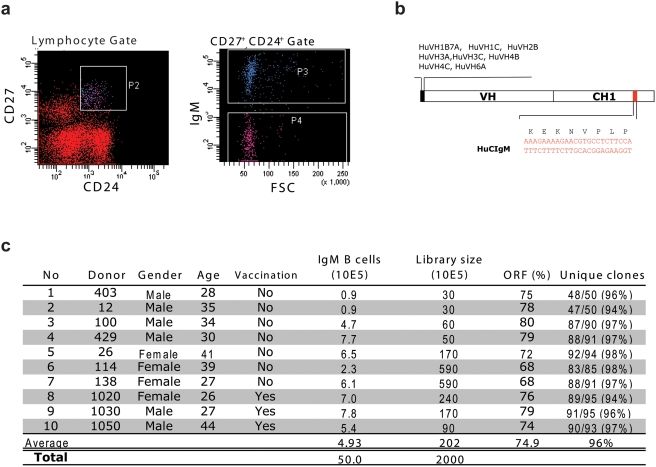
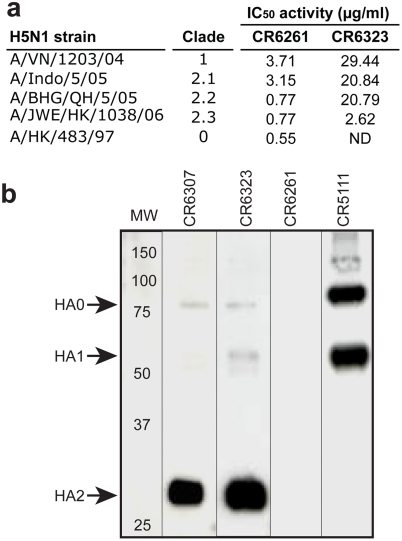
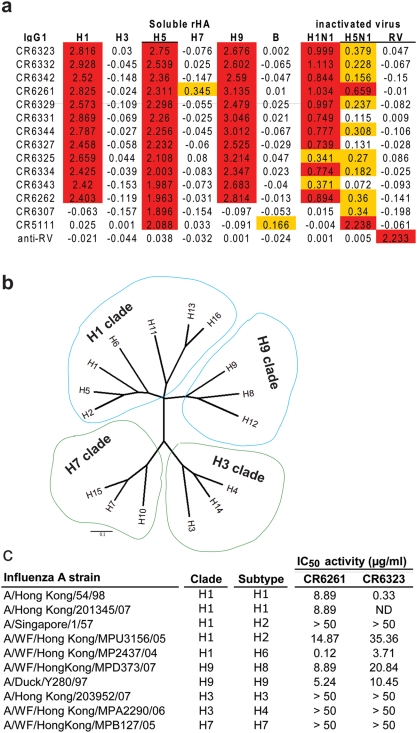
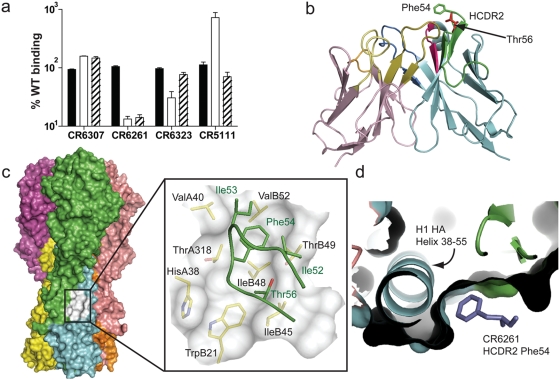
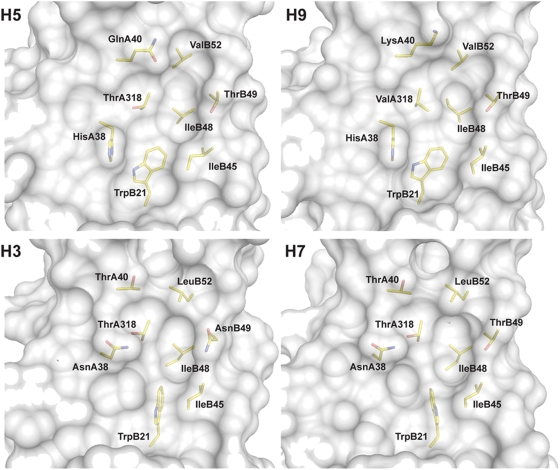
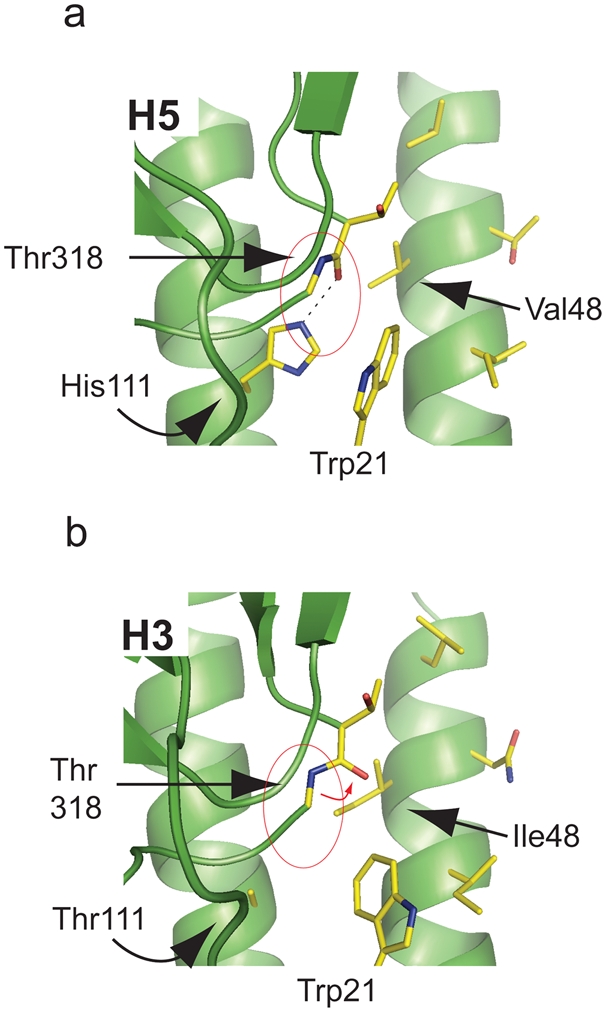
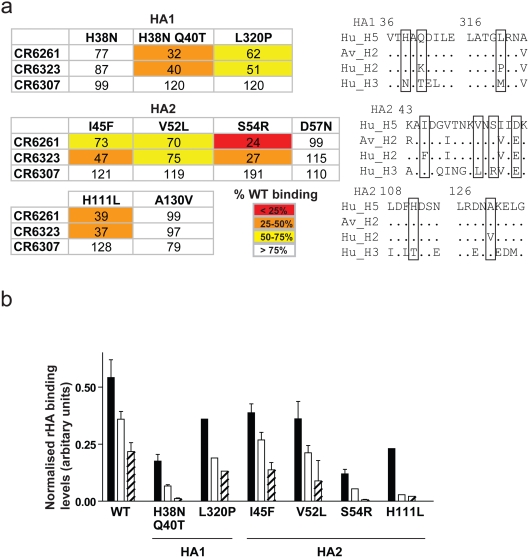
Methods and findings: Here we describe a panel of 13 monoclonal antibodies (mAbs) recovered from combinatorial display libraries that were constructed from human IgM(+) memory B cells of recent (seasonal) influenza vaccinees. The mAbs have broad heterosubtypic neutralizing activity against antigenically diverse H1, H2, H5, H6, H8 and H9 influenza subtypes. Restriction to variable heavy chain gene IGHV1-69 in the high affinity mAb panel was associated with binding to a conserved hydrophobic pocket in the stem domain of HA. The most potent antibody (CR6261) was protective in mice when given before and after lethal H5N1 or H1N1 challenge.
Conclusions: The human monoclonal CR6261 described in this study could be developed for use as a broad spectrum agent for prophylaxis or treatment of human or avian influenza infections without prior strain characterization. Moreover, the CR6261 epitope could be applied in targeted vaccine strategies or in the design of novel antivirals. Finally our approach of screening the IgM(+) memory repertoire could be applied to identify conserved and functionally relevant targets on other rapidly evolving pathogens.
Conflict of interest statement
Figures


References
-
- WHO. Cumulative Number of Confirmed Human Cases of Avian Influenza A/(H5N1) Reported to WHO. 2008 http://www.who.int/csr/disease/avian_influenza/country/cases_table_2008_....
-
- Peiris M, Yuen KY, Leung CW, Chan KH, Ip PL, et al. Human infection with influenza H9N2. Lancet. 1999;354:916–917. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
