

Structural basis for inactivation of the human pyruvate dehydrogenase complex by phosphorylation: role of disordered phosphorylation loops
- PMID: 19081061
- PMCID: PMC2849990
- DOI: 10.1016/j.str.2008.10.010
Structural basis for inactivation of the human pyruvate dehydrogenase complex by phosphorylation: role of disordered phosphorylation loops
Abstract
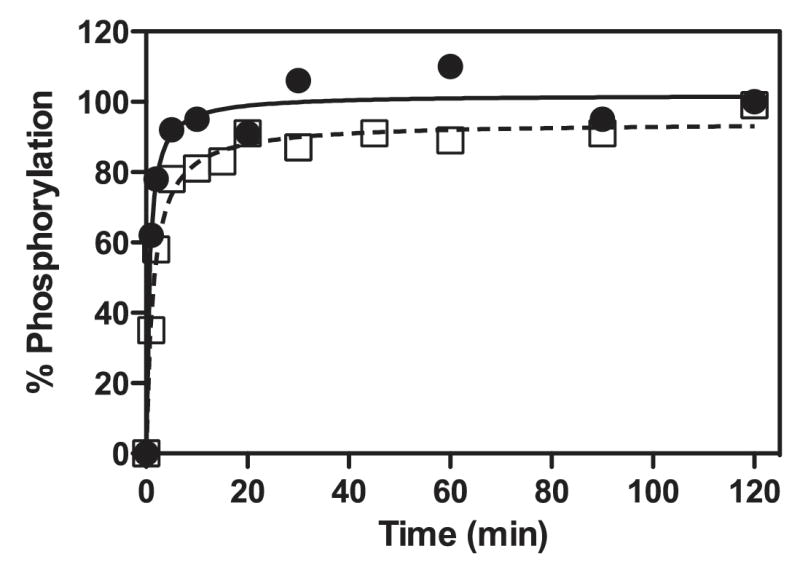
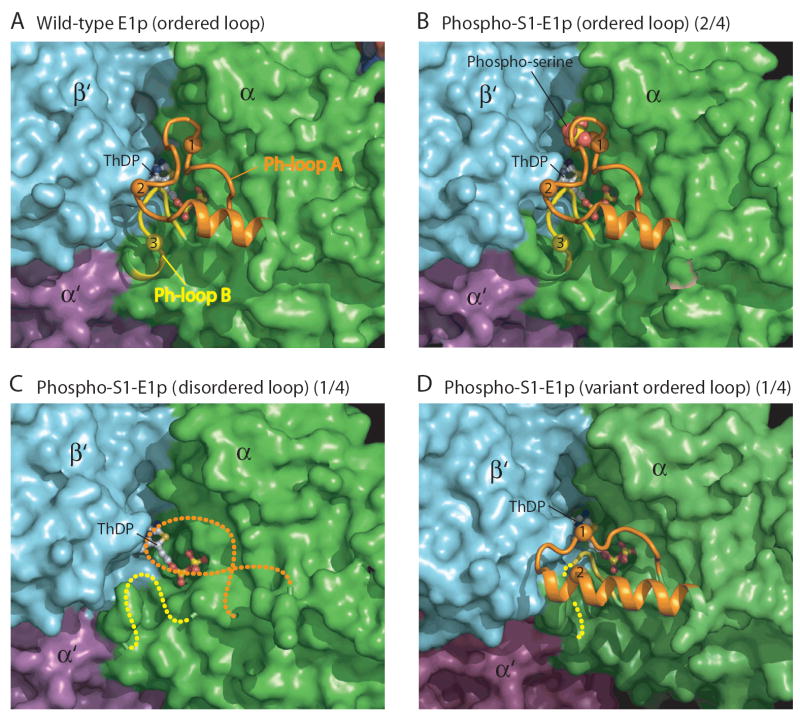
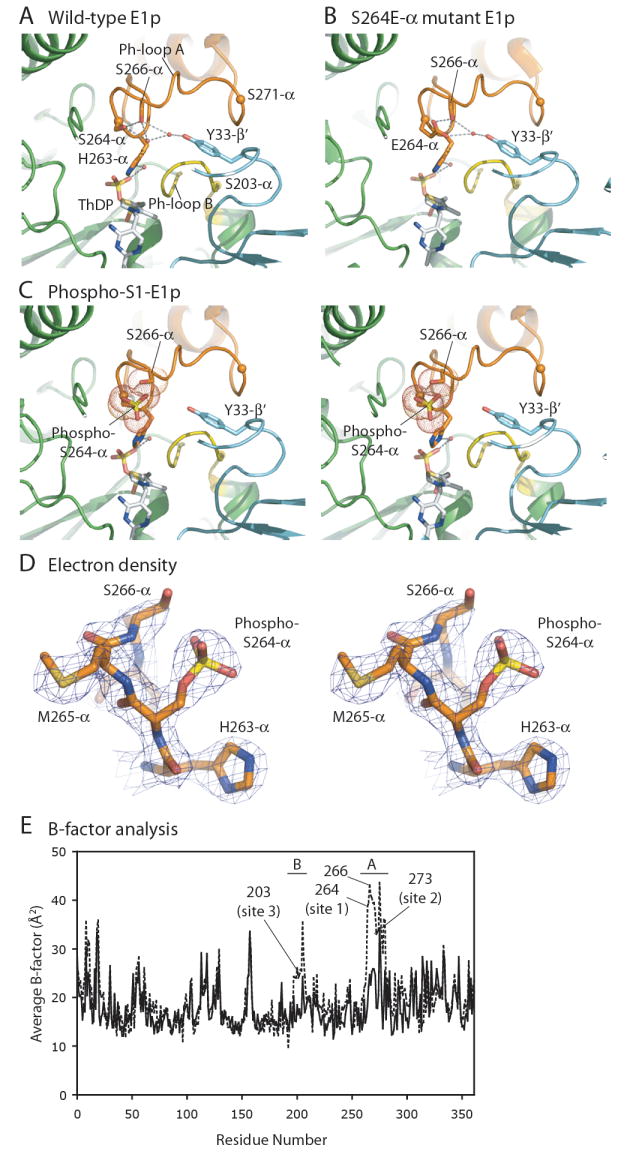
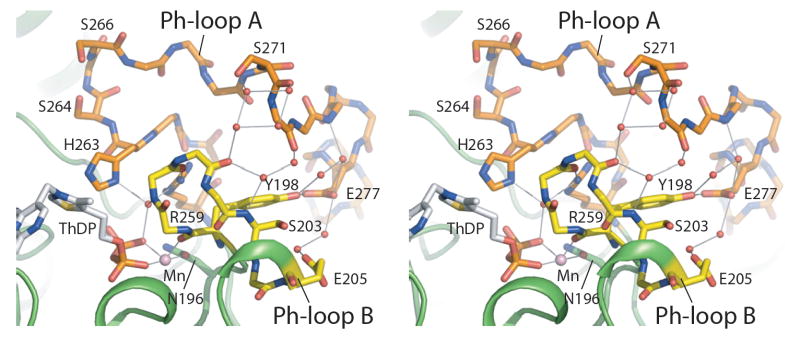
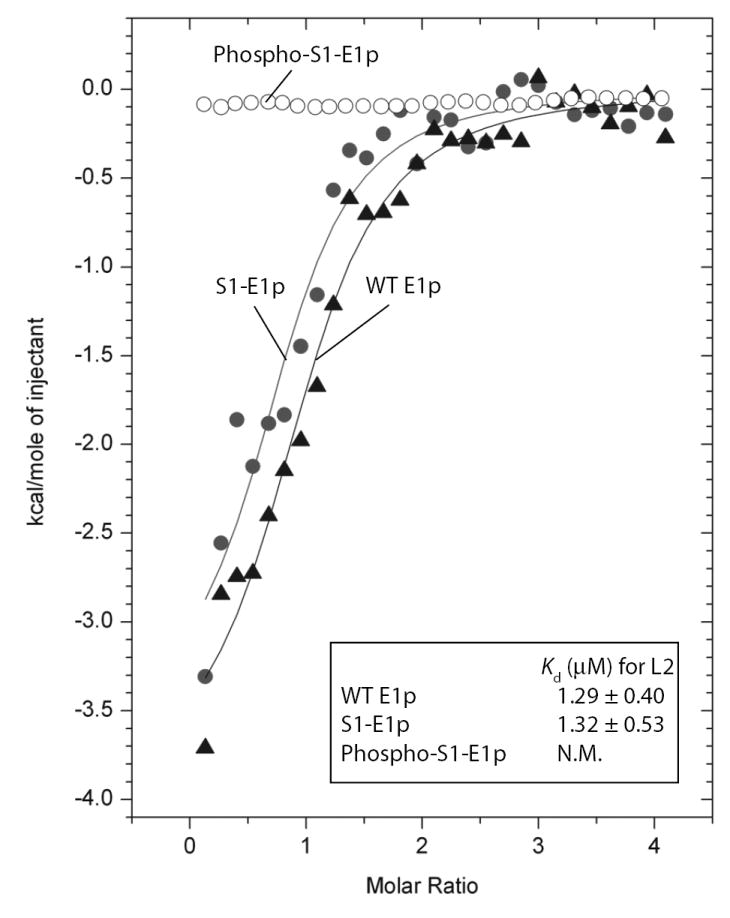
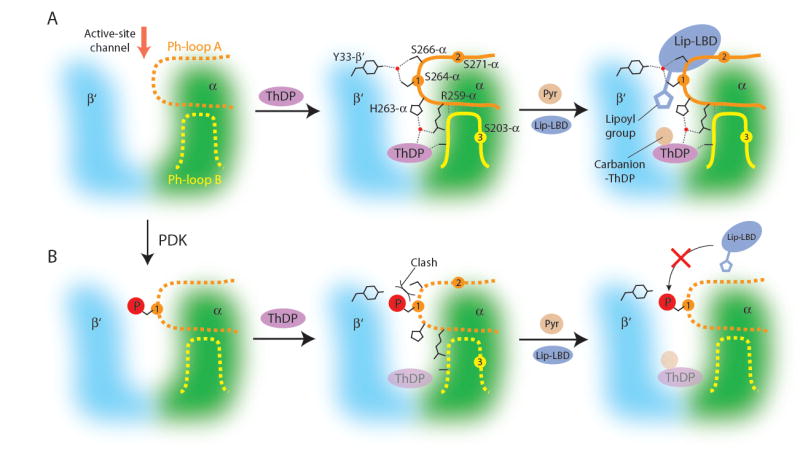
We report the crystal structures of the phosporylated pyruvate dehydrogenase (E1p) component of the human pyruvate dehydrogenase complex (PDC). The complete phosphorylation at Ser264-alpha (site 1) of a variant E1p protein was achieved using robust pyruvate dehydrogenase kinase 4 free of the PDC core. We show that unlike its unmodified counterpart, the presence of a phosphoryl group at Ser264-alpha prevents the cofactor thiamine diphosphate-induced ordering of the two loops carrying the three phosphorylation sites. The disordering of these phosphorylation loops is caused by a previously unrecognized steric clash between the phosphoryl group at site 1 and a nearby Ser266-alpha, which nullifies a hydrogen-bonding network essential for maintaining the loop conformations. The disordered phosphorylation loops impede the binding of lipoyl domains of the PDC core to E1p, negating the reductive acetylation step. This results in the disruption of the substrate channeling in the PDC, leading to the inactivation of this catalytic machine.
Conflict of interest statement
The authors declare no financial conflict of interest related to this work.
Figures

References
-
- Ali MS, Shenoy BC, Eswaran D, Andersson LA, Roche TE, Patel MS. Identification of the tryptophan residue in the thiamin pyrophosphate binding site of mammalian pyruvate dehydrogenase. J Biol Chem. 1995;270:4570–4574. - PubMed
-
- Bao H, Kasten SA, Yan X, Hiromasa Y, Roche TE. Pyruvate Dehydrogenase Kinase Isoform 2 Activity Stimulated by Speeding Up the Rate of Dissociation of ADP. Biochemistry. 2004;43:13442–13451. - PubMed
-
- Bonnet S, Archer SL, Allalunis-Turner J, Haromy A, Beaulieu C, Thompson R, Lee CT, Lopaschuk GD, Puttagunta L, Bonnet S, et al. A mitochondria-K+ channel axis is suppressed in cancer and its normalization promotes apoptosis and inhibits cancer growth. Cancer cell. 2007;11:37–51. - PubMed
-
- CCP4. The CCP4 suite: programs for protein crystallography. Acta Crystallogr D Biol Crystallogr. 1994;50:760–763. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
