

Review
doi: 10.1016/j.devcel.2008.11.010.
Hedgehog signaling in development and cancer
Affiliations
- PMID: 19081070
- PMCID: PMC6443374
- DOI: 10.1016/j.devcel.2008.11.010
Item in Clipboard
Review
Hedgehog signaling in development and cancer
Dev Cell.
2008 Dec.
Abstract
The Hedgehog (Hh) family of secreted proteins governs a wide variety of processes during embryonic development and adult tissue homeostasis. Here we review the current understanding of the molecular and cellular basis of Hh morphogen gradient formation and signal transduction, and the multifaceted roles of Hh signaling in development and tumorigenesis. We discuss how the Hh pathway has diverged during evolution and how it integrates with other signaling pathways to control cell growth and patterning.
Figures
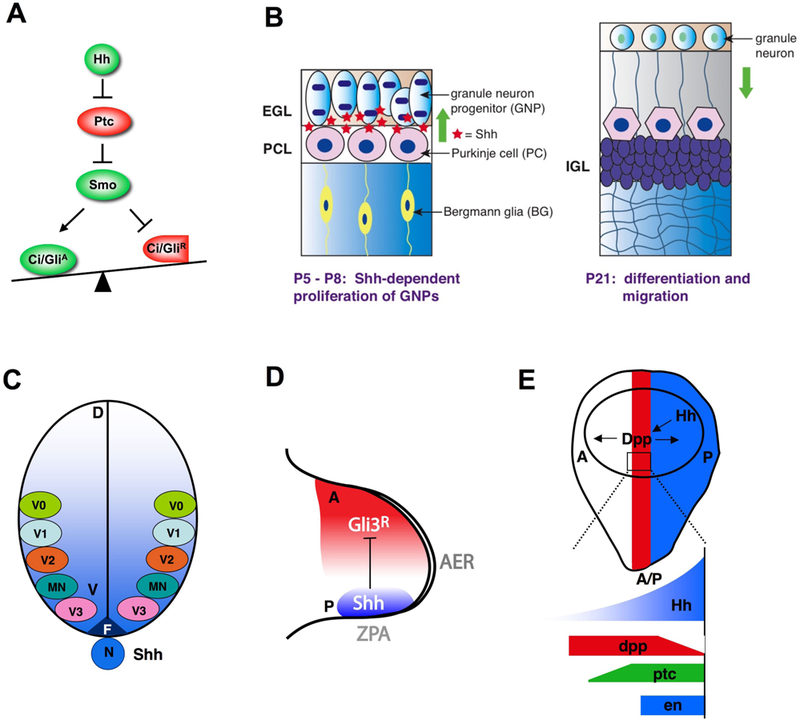
(A) A simplified Hh signaling pathway. (B) In cerebellar development, Shh secreted by Purkinje cells controls the proliferation of granule neuron progenitors in the external germinal cell layer (EGL). (C) In developing neural tube, Shh emanates from notochord (N) and floor-plate (F) to form a ventral (V) to dorsal (D) concentration gradient that directs the formation of distinct pools of neural progenitor cells at different positions along the D/V axis. Increasing levels of Shh progressively specify progenitors giving rise to neuronal subtypes of more ventral characters. MN: motor neuron; V0-V3: V0-V3 interneurons. (D) In vertebrate limb bud, Shh produced by the ZPA propagates anteriorly to form a posterior to anterior concentration gradient that patterns the limb. (E) In Drosophila wing imaginal disc, Hh produced by posterior (P) compartment cells (blue) acts as a local morphogen, inducing gene expression in the neighboring anterior (A) compartment cells (red). Low levels of Hh suffice to activate dpp while higher levels of Hh are required to activate ptc, and peak levels of Hh are required to activate en.
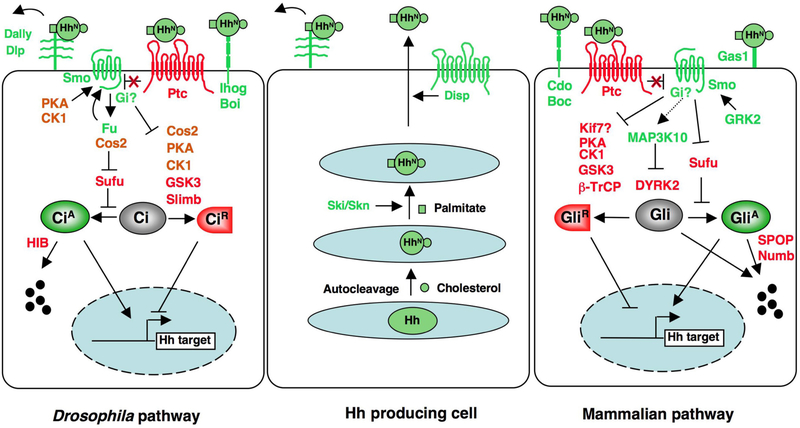
In Hh-producing cells, full-length Hh is auto-catalytically cleaved to generate an N-terminal fragment (HhN) modified by cholesterol. HhN is palmitoylated by Ski/Skn. Secretion of dual lipid modified Hh is mediated by Disp. Movement of Hh requires the HSPGs, Dally and Dlp, in Drosophila. Hh signal reception is facilitated by Ihog/Boi in Drosophila and Cdo/Boc/Gas1 in mammals. In the absence of Hh, Ptc blocks Smo activation and full-length Ci/Gli (Gli2 and Gli3) is phosphorylated by multiple kinases and subsequently targeted to ubiquitin/proteasome-mediated proteolysis through SCFSlimb/β-TRCP to generate a truncated repressor form (CiR/GliR). In Drosophila, efficient phosphorylation of Ci requires the kinesin like protein Cos2, which acts as a molecular scaffold to bridge Ci and its kinases. Hh binding to Ptc blocks its inhibition of Smo. In Drosophila, this triggers Smo phosphorylation by PKA and CKI, leading to the cell surface accumulation and activation of Smo. Smo then recruits Cos2-Fu to activate Fu and dissociates Cos2-Ci-kinase complexes to inhibit Ci phosphorylation and processing. Furthermore, high levels of Hh stimulate CiA via Fu-mediated antagonism of Sufu. Hh signaling induces the expression of nuclear HIB that targets CiA for degradation. Fu-Cos2 is also involved in a feedback regulation of Smo phosphorylation. In mammalian systems, Fu homolog is not required for Hh signaling and the involvement of Cos2 homologs, such as Kif7, is uncertain. Sufu is the major Gli inhibitor. DYRK2 phosphorylates Gli2 and targets it for degradation whereas MAP3K10 activates Gli2 in part by inhibiting DYKR2. Smo phosphorylation requires GRK2. Gαi has been implicated in Hh signaling downstream of Smo in both Drosophila and vertebrates.
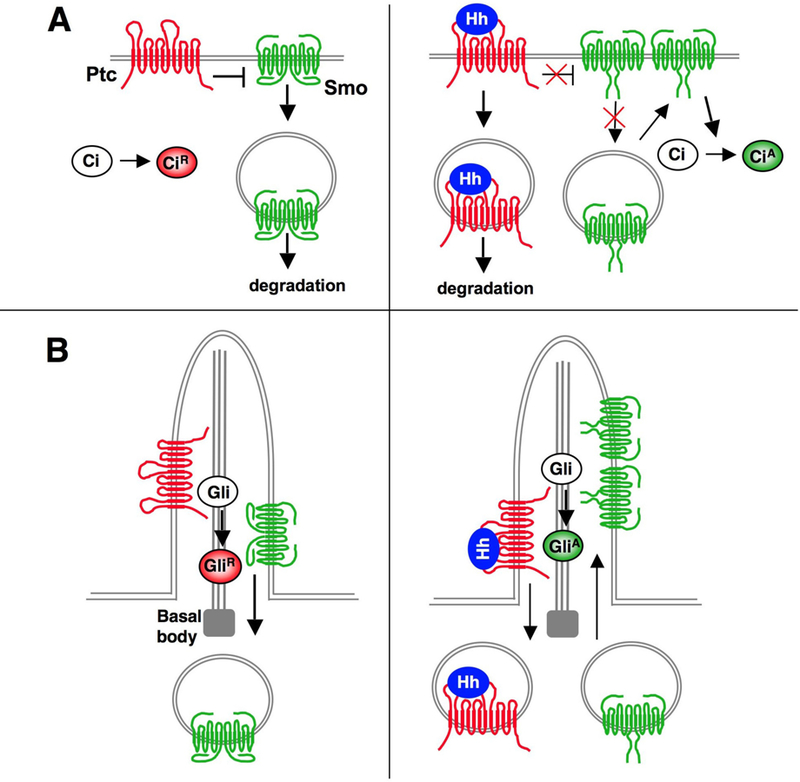
(A) In Drosophila, the absence of Hh allows Ptc to inhibit Smo phosphorylation, which promotes Smo endocytosis and degradation. Binding of Hh to Ptc stimulates Smo hyperphosphorylation, which increases Smo cell surface expression by inhibiting endocytosis and/or promoting recycling. In addition, phosphorylation of Smo promotes a conformational change leading to oligomerization of Smo C-tails (B) In mammals, Ptc resides in the primary cilium and prevents Smo ciliary accumulation in the absence of Hh. Hh binding to Ptc promotes the exit of Ptc from the cilium and allows Smo to accumulate in the cilium. Hh also induces conformational change and clustering of Smo C-tails.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
