

Inhibitors of melanogenesis increase toxicity of cyclophosphamide and lymphocytes against melanoma cells
- PMID: 19085934
- PMCID: PMC2628959
- DOI: 10.1002/ijc.24005
Inhibitors of melanogenesis increase toxicity of cyclophosphamide and lymphocytes against melanoma cells
Abstract
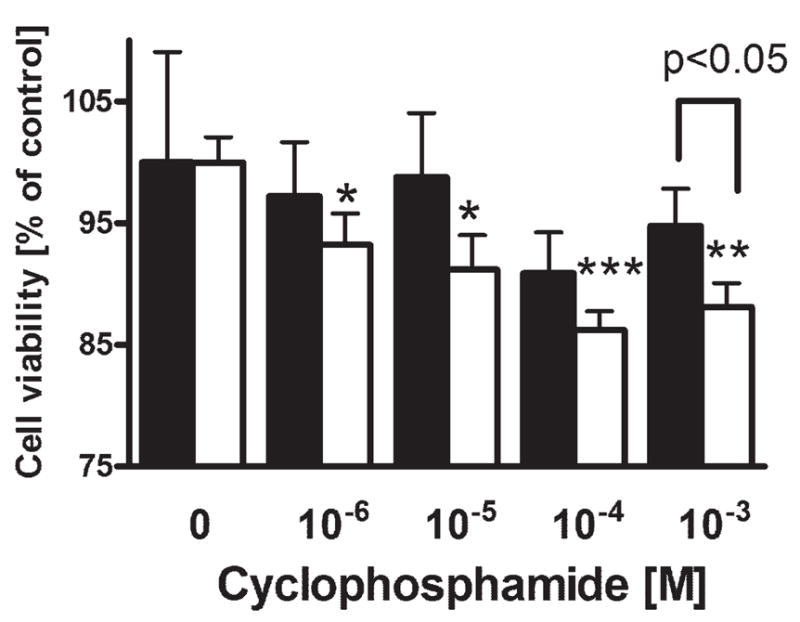
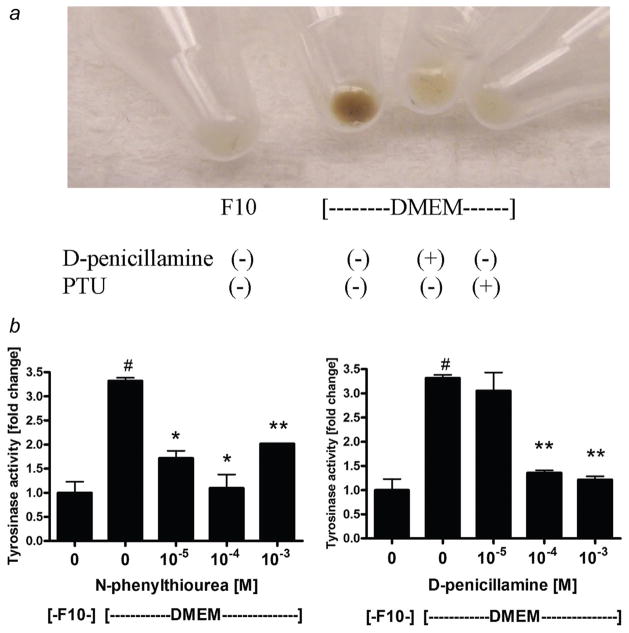
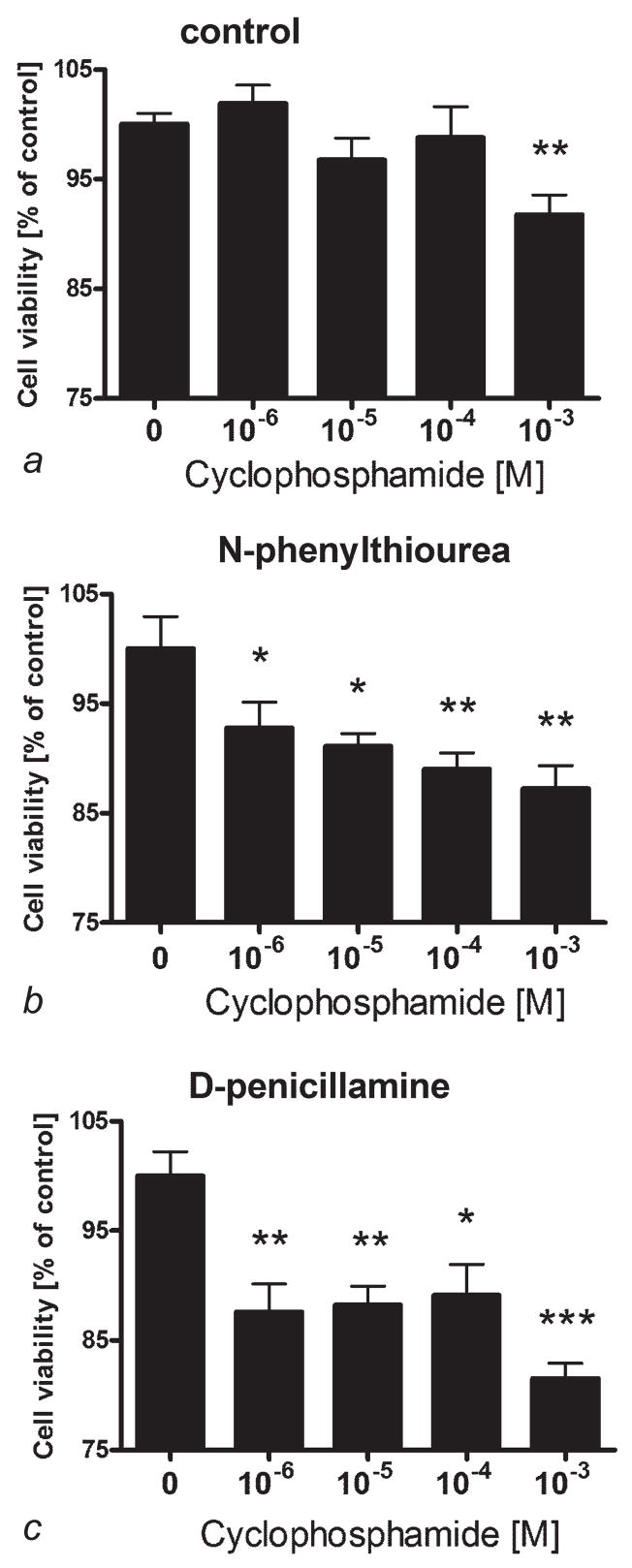
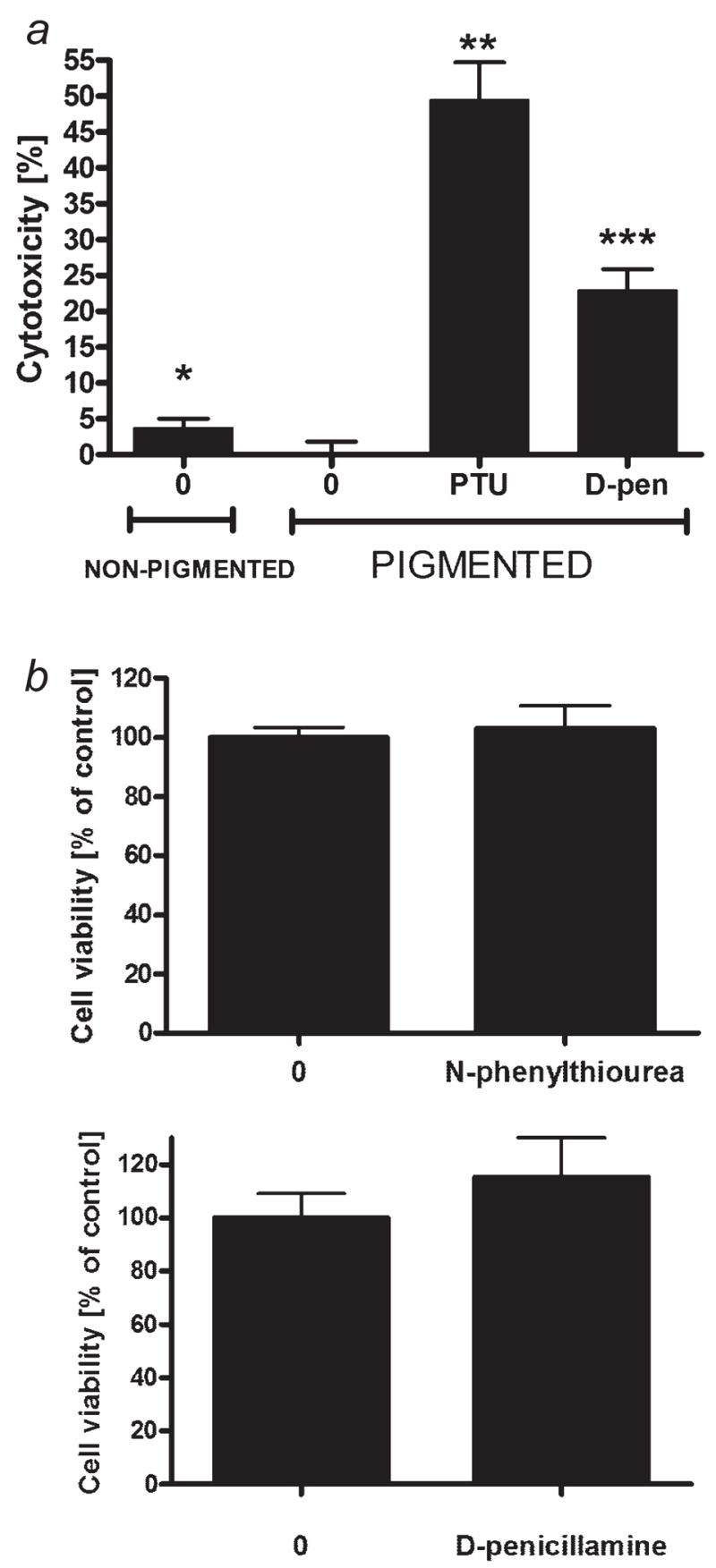
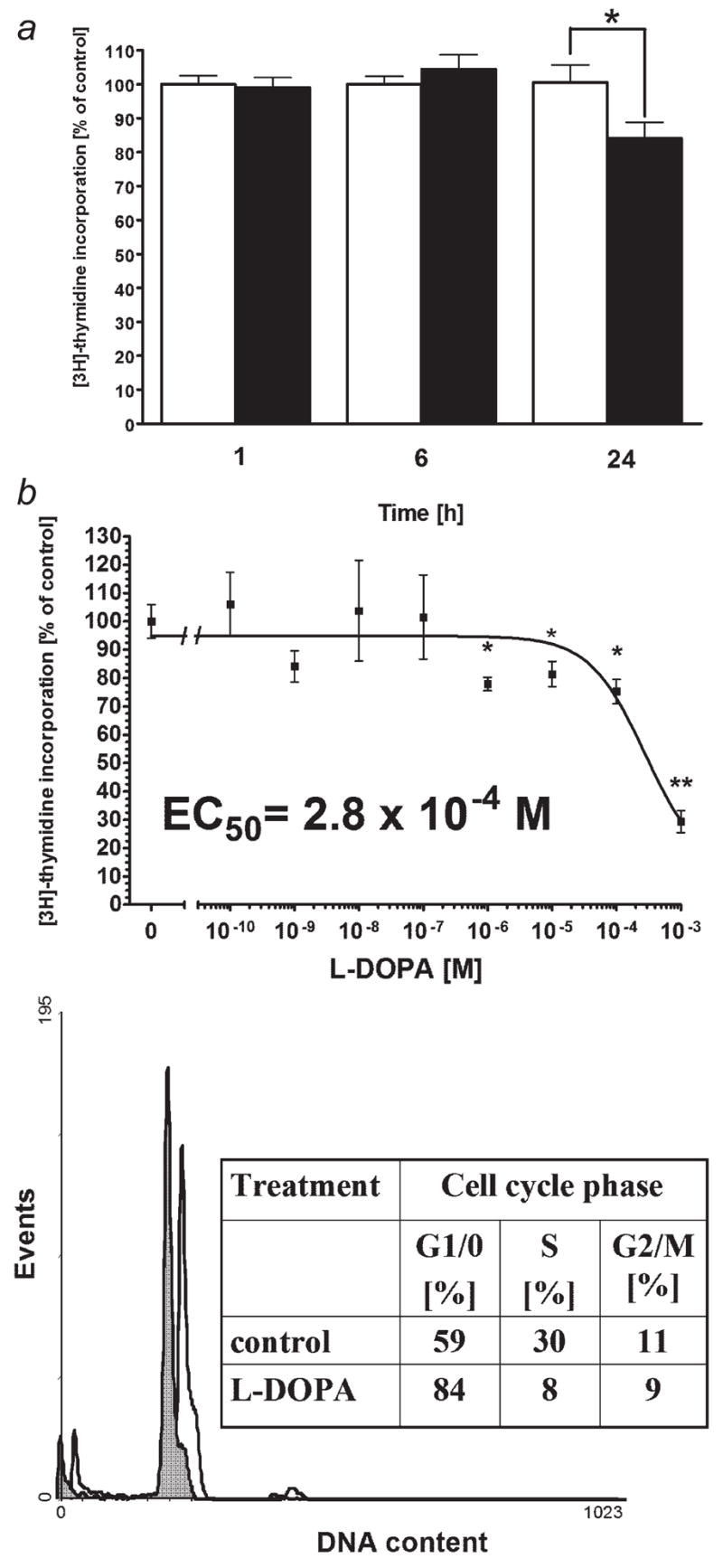
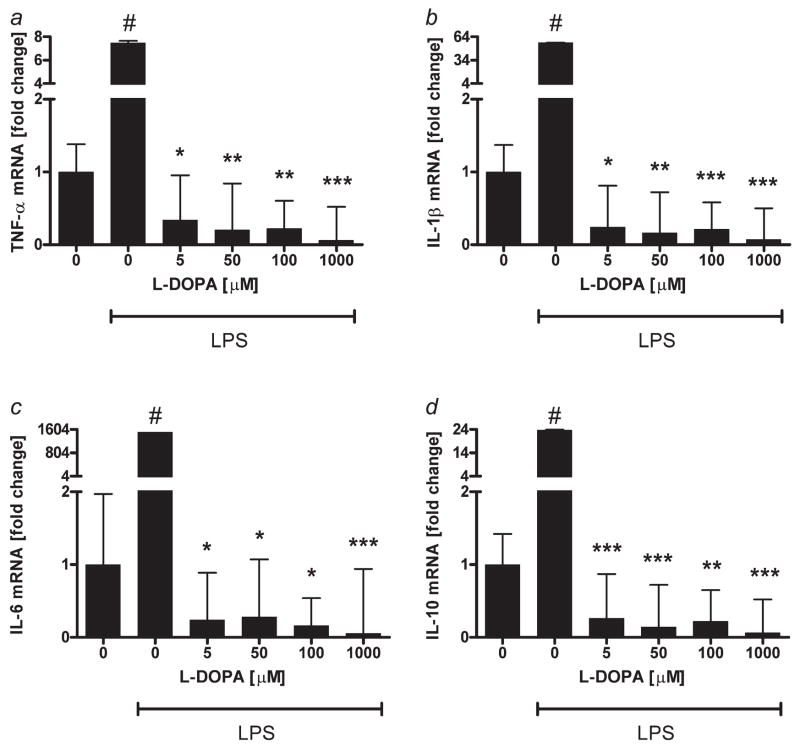
High mortality rate for metastatic melanoma is related to its resistant to the current methods of therapy. Melanogenesis is a metabolic pathway characteristic for normal and malignant melanocytes that can affect the behavior of melanoma cells or its surrounding environment. Human melanoma cells in which production of melanin pigment is dependent on tyrosine levels in medium were used for experiments. Peripheral blood mononuclear cells were derived from the buffy coats purchased from Lifeblood Biological Services. Cell pigmentation was evaluated macroscopically, and tyrosinase activity was measured spectrophotometrically. Cell proliferation and viability were measured using lactate dehydrogenase release MTT, [(3)H]-thymidine incorporation and DNA content analyses, and gene expression was measured by real time RT-PCR. Pigmented melanoma cells were significantly less sensitive to cyclophosphamide and to killing action of IL-2-activated peripheral blood lymphocytes. The inhibition of melanogenesis by either blocking tyrosinase catalytic site or chelating copper ions sensitized melanoma cells towards cytotoxic action of cyclophosphamide, and amplified immunotoxic activities of IL-2 activated lymphocytes. Exogenous L-DOPA inhibited lymphocyte proliferation producing the cell cycle arrest in G1/0 and dramatically inhibited the production of IL-1beta, TNF-alpha, IL-6 and IL-10. Thus, the active melanogenesis could not only impair the cytotoxic action of cyclophosphamid but also has potent immunosuppressive properties. This resistance to a chemotherapeutic agent or immunotoxic activity of lymphocytes could be reverted by the action of tyrosinase inhibitors. Thus, the inhibition of melanogenesis might represent a valid therapeutic target for the management of advanced melanotic melanomas.
Figures
References
-
- Miller AJ, Mihm MC., Jr Melanoma. N Engl J Med. 2006;355:51–65. - PubMed
-
- Becker D, Mihm MC, Hewitt SM, Sondak VK, Fountain JW, Thurin M. Markers and tissue resources for melanoma: meeting report. Cancer Res. 2006;66:10652–7. - PubMed
-
- Carlson JA, Ross JS, Slominski A, Linette G, Mysliborski J, Hill J, Mihm M., Jr Molecular diagnostics in melanoma. J Am Acad Dermatol. 2005;52:743–75. quiz 75–8. - PubMed
-
- Meyskens FL, Jr, Farmer PJ, Anton-Culver H. Etiologic pathogenesis of melanoma: a unifying hypothesis for the missing attributable risk. Clin Cancer Res. 2004;10:2581–3. - PubMed
-
- Markovic SN, Erickson LA, Rao RD, Weenig RH, Pockaj BA, Bardia A, Vachon CM, Schild SE, McWilliams RR, Hand JL, Laman SD, Kottschade LA, et al. Malignant melanoma in the 21st century, part 1: epidemiology, risk factors, screening, prevention, and diagnosis. Mayo Clin Proc. 2007;82:364–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
