

TGF-beta promotes cell death and suppresses lactation during the second stage of mammary involution
- PMID: 19086032
- PMCID: PMC3038423
- DOI: 10.1002/jcp.21646
TGF-beta promotes cell death and suppresses lactation during the second stage of mammary involution
Abstract
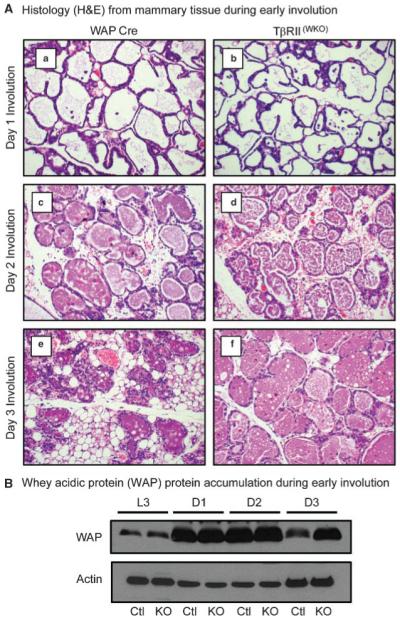
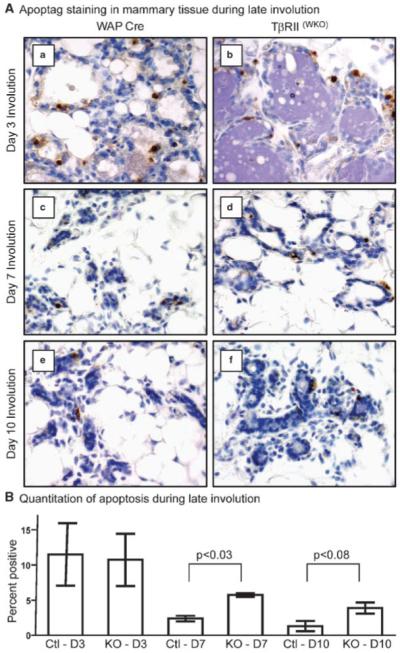
Transforming growth factor beta (TGF-beta) ligands are known to regulate virgin mammary development and contribute to initiation of post-lactation involution. However, the role for TGF-beta during the second phase of mammary involution has not been addressed. Previously, we have used an MMTV-Cre transgene to delete exon 2 from the Tgfbr2 gene in mammary epithelium, however we observed a gradual loss of T beta RII deficient epithelial cells that precluded an accurate study of the role for TGF-beta signaling during involution timepoints. Therefore, in order to determine the role for TGF-beta during the second phase of mammary involution we have now targeted T beta RII ablation within mammary epithelium using the WAP-Cre transgene [T beta RII(WKO)Rosa26R]. Our results demonstrated that TGF-beta regulates commitment to cell death during the second phase of mammary involution. Importantly, at day 3 of mammary involution the Na-Pi type IIb co-transporter (Npt2b), a selective marker for active lactation in luminal lobular alveolar epithelium, was completely silenced in the WAP-Cre control and T beta RII(WKO)Rosa26R tissues. However, by day 7 of involution the T beta RII(WKO)Rosa26R tissues had distended lobular alveoli and regained a robust Npt2b signal that was detected at the apical luminal surface. The Npt2b abundance and localization positively correlated with elevated WAP mRNA expression, suggesting that the distended alveoli were the result of an active lactation program rather than residual milk protein and lipid accumulation. In summary, the results suggest that an epithelial cell response to TGF-beta signaling regulates commitment to cell death and suppression of lactation during the second phase of mammary involution.
(c) 2008 Wiley-Liss, Inc.
Figures




References
-
- Bailey JP, Nieport KM, Herbst MP, Srivastava S, Serra RA, Horseman ND. Prolactin and transforming growth factor-beta signaling exert opposing effects on mammary gland morphogenesis, involution, and the Akt-forkhead pathway. Mol Endocrinol (Baltimore, MD) 2004;18:1171–1184. - PubMed
-
- Bierie B, Stover DG, Abel TW, Chytil A, Gorska AE, Aakre M, Forrester E, Yang L, Wagner KU, Moses HL. Transforming growth factor-beta regulates mammary carcinoma cell survival and interaction with the adjacent microenvironment. Cancer Res. 2008;68:1809–1819. - PubMed
-
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
