

Bursting activation of prefrontal cortex drives sustained up states in nucleus accumbens spiny neurons in vivo
- PMID: 19086088
- PMCID: PMC2628410
- DOI: 10.1002/syn.20593
Bursting activation of prefrontal cortex drives sustained up states in nucleus accumbens spiny neurons in vivo
Abstract
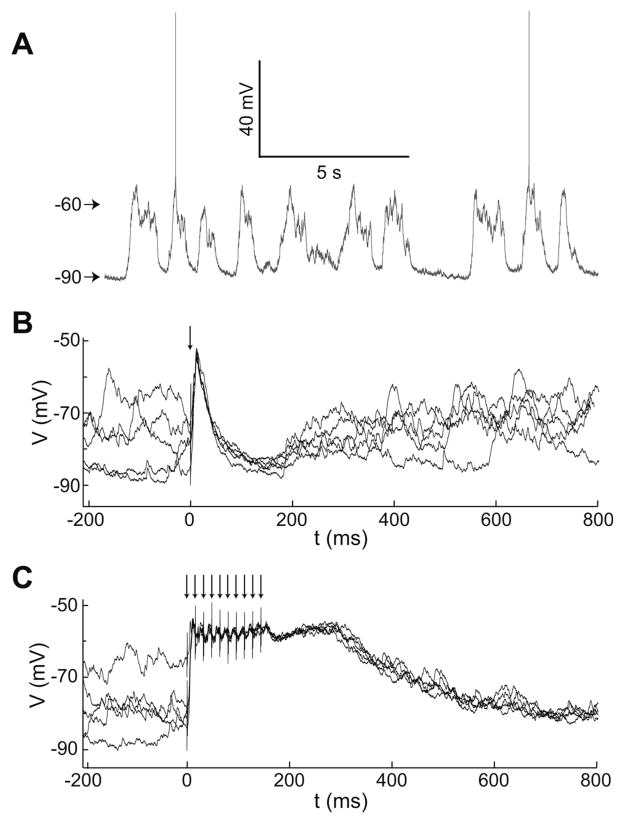
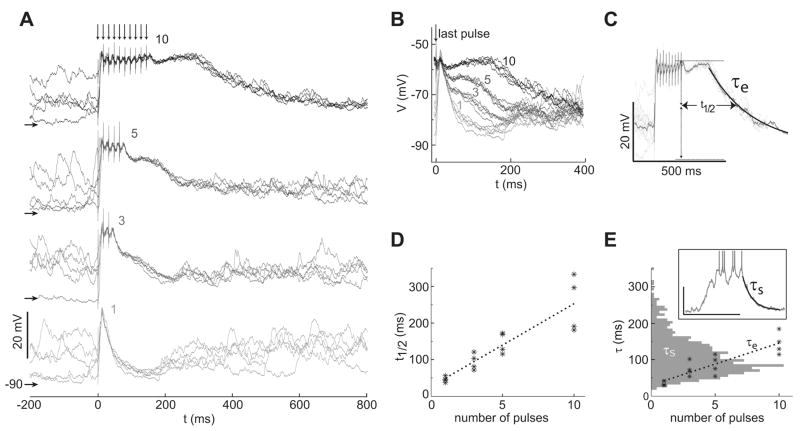
Hippocampal inputs to the nucleus accumbens (NA) have been proposed to implement a gating mechanism by driving NA medium spiny neurons (MSNs) to depolarized up states that facilitate action potential firing in response to brief activation of the prefrontal cortex (PFC). Brief PFC stimulation alone, on the other hand, could not drive NA up states. As these studies were conducted using single-pulse PFC stimulation, it remains possible that PFC activation with naturalistic, bursty patterns can also drive up states in NA MSNs. Here, we assessed NA responses to PFC stimulation with a pattern similar to what is typically observed in awake animals during PFC-relevant behaviors. In vivo intracellular recordings from NA MSNs revealed that brief 20-50 Hz PFC stimulus trains evoked depolarizations that were similar to spontaneous up states in NA MSNs and were sustained beyond stimulus offset. Similar train stimulation of corticoaccumbens afferents in a parasagittal slice preparation evoked large amplitude depolarizations in NA MSNs that were sustained during stimulation but decayed rapidly following stimulation offset, suggesting that activation of cortical afferents can drive MSN depolarizations but other mechanisms may contribute to sustaining up states. These data suggest that NA MSNs integrate temporal features of PFC activation and that the NA gating model can be reformulated to include a PFC-driven gating mechanism during periods of high PFC firing, such as during cognitively demanding tasks. Synapse 63:173-180, 2009. (c) 2008 Wiley-Liss, Inc.
Figures


References
-
- Block AE, Dhanji H, Thompson-Tardif SF, Floresco SB. Thalamic-prefrontal cortical-ventral striatal circuitry mediates dissociable components of strategy set shifting. Cereb Cortex. 2007;17:1625–1636. - PubMed
-
- Calabresi P, Pisani A, Mercuri NB, Bernardi G. Long-term potentiation in the striatum is unmasked by removing the voltage-dependent magnesium block of NMDA receptor channels. Eur J Neurosci. 1992;4:929–935. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous