

The Bax/Bak ortholog in Drosophila, Debcl, exerts limited control over programmed cell death
- PMID: 19088092
- PMCID: PMC2685970
- DOI: 10.1242/dev.019042
The Bax/Bak ortholog in Drosophila, Debcl, exerts limited control over programmed cell death
Abstract
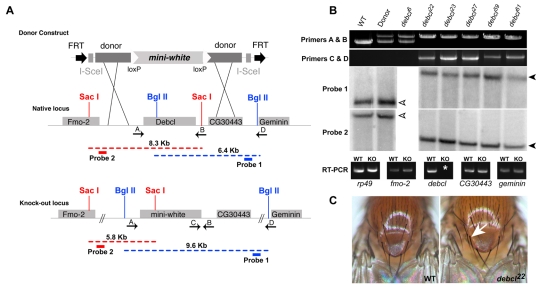
Bcl-2 family members are pivotal regulators of programmed cell death (PCD). In mammals, pro-apoptotic Bcl-2 family members initiate early apoptotic signals by causing the release of cytochrome c from the mitochondria, a step necessary for the initiation of the caspase cascade. Worms and flies do not show a requirement for cytochrome c during apoptosis, but both model systems express pro- and anti-apoptotic Bcl-2 family members. Drosophila encodes two Bcl-2 family members, Debcl (pro-apoptotic) and Buffy (anti-apoptotic). To understand the role of Debcl in Drosophila apoptosis, we produced authentic null alleles at this locus. Although gross development and lifespans were unaffected, we found that Debcl was required for pruning cells in the developing central nervous system. debcl genetically interacted with the ced-4/Apaf1 counterpart dark, but was not required for killing by RHG (Reaper, Hid, Grim) proteins. We found that debcl(KO) mutants were unaffected for mitochondrial density or volume but, surprisingly, in a model of caspase-independent cell death, heterologous killing by murine Bax required debcl to exert its pro-apoptotic activity. Therefore, although debcl functions as a limited effector of PCD during normal Drosophila development, it can be effectively recruited for killing by mammalian members of the Bcl-2 gene family.
Figures






References
-
- Abdelwahid, E., Yokokura, T., Krieser, R. J., Balasundaram, S., Fowle, W. H. and White, K. (2007). Mitochondrial disruption in Drosophila apoptosis. Dev. Cell 12, 793-806. - PubMed
-
- Arama, E., Agapite, J. and Steller, H. (2003). Caspase activity and a specific cytochrome C are required for sperm differentiation in Drosophila. Dev. Cell 4, 687-697. - PubMed
-
- Brachmann, C. B., Jassim, O. W., Wachsmuth, B. D. and Cagan, R. L. (2000). The Drosophila Bcl-2 family member dBorg-1 functions in the apoptotic response to UV-irradiation. Curr. Biol. 10, 547-550. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
