

E3 ligase activity of BRCA1 is not essential for mammalian cell viability or homology-directed repair of double-strand DNA breaks
- PMID: 19088202
- PMCID: PMC2603436
- DOI: 10.1073/pnas.0811203106
E3 ligase activity of BRCA1 is not essential for mammalian cell viability or homology-directed repair of double-strand DNA breaks
Abstract
Hereditary cases of breast and ovarian cancer are often attributed to germ-line mutations of the BRCA1 tumor suppressor gene. Although BRCA1 is involved in diverse cellular processes, its role in the maintenance of genomic integrity may be a key component of its tumor suppression activity. The protein encoded by BRCA1 interacts in vivo with the related BARD1 protein to form a heterodimeric complex that acts as a ubiquitin E3 ligase. Because the enzymatic activity of the BRCA1/BARD1 heterodimer is conserved over a broad phylogenetic range, it is thought to be critical for the central functions of BRCA1. To test this hypothesis, we have generated isogenic clones of embryonic stem cells that do or do not express an enzymatically proficient Brca1 polypeptide. Surprisingly, cells lacking the ubiquitin ligase activity of BRCA1 are viable and do not accumulate spontaneous cytogenetic rearrangements. Gene targeting efficiencies are modestly reduced in these cells, and chromosomal rearrangements arise at elevated rates in response to genotoxic stress. Nonetheless, cells lacking Brca1 enzymatic activity are not hypersensitive to the DNA cross-linking agent mitomycin C. They also form Rad51 focus in response to ionizing radiation and repair chromosome breaks by homologous recombination at wild-type levels. These results indicate that key aspects of BRCA1 function in genome maintenance, including its role in homology-directed repair of double-strand DNA breaks, do not depend on the E3 ligase activity of BRCA1.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
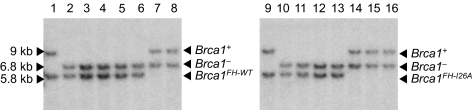
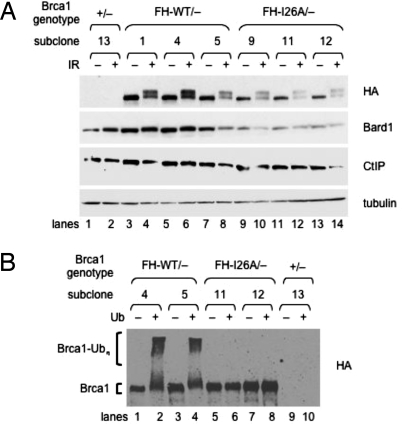
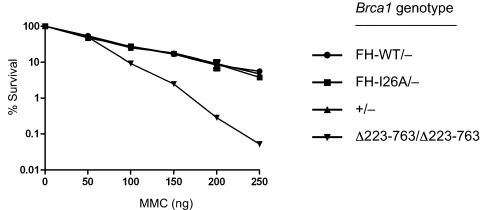
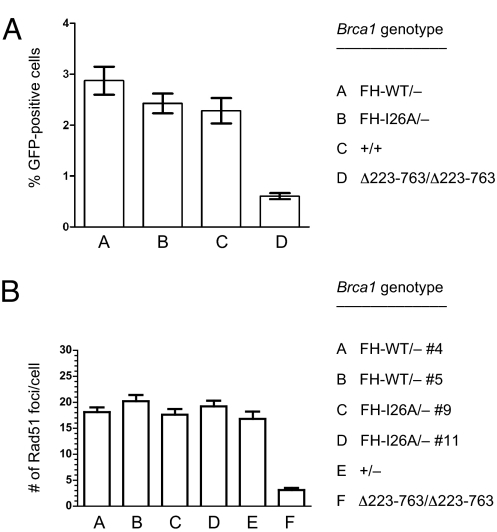
References
-
- Wooster R, Weber BL. Breast and ovarian cancer. N Engl J Med. 2003;348:2339–2347. - PubMed
-
- Evers B, Jonkers J. Mouse models of BRCA1 and BRCA2 deficiency: Past lessons, current understanding and future prospects. Oncogene. 2006;25:5885–5897. - PubMed
-
- Moynahan ME, Cui TY, Jasin M. Homology-directed DNA repair, mitomycin-c resistance, and chromosome stability is restored with correction of a Brca1 mutation. Cancer Res. 2001;61:4842–4850. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
