

Genome-wide profiling of salt fractions maps physical properties of chromatin
- PMID: 19088306
- PMCID: PMC2661814
- DOI: 10.1101/gr.087619.108
Genome-wide profiling of salt fractions maps physical properties of chromatin
Abstract
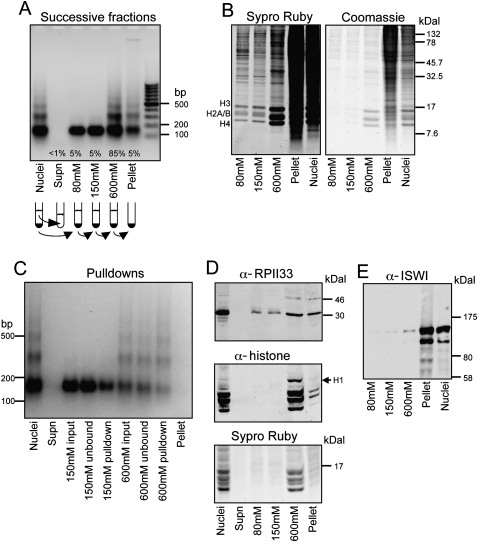
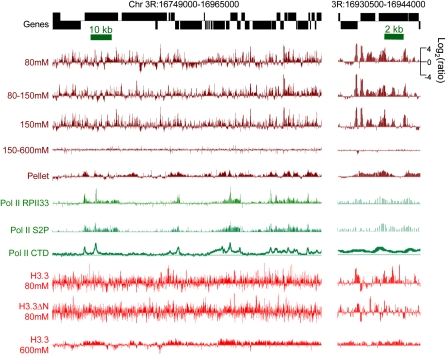
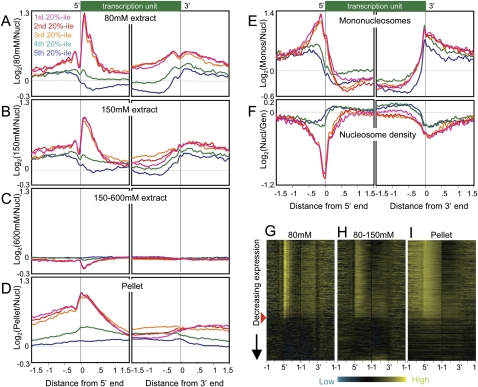
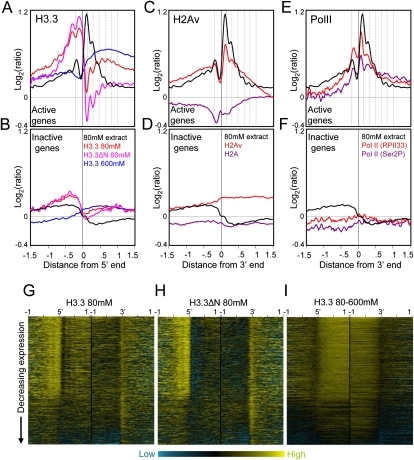
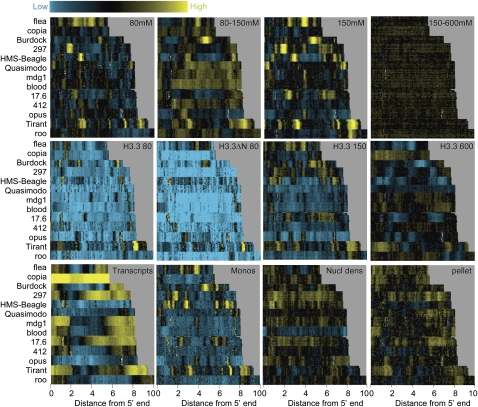
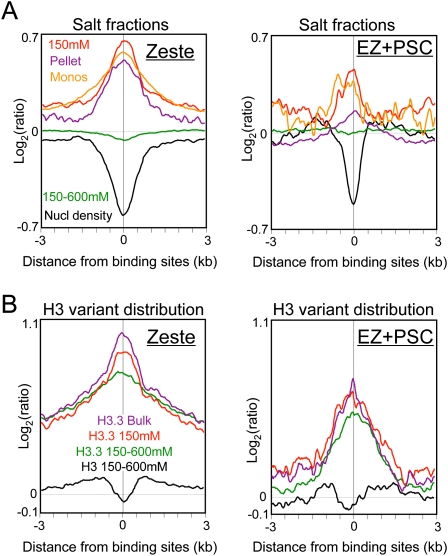
We applied genome-wide profiling to successive salt-extracted fractions of micrococcal nuclease-treated Drosophila chromatin. Chromatin fractions extracted with 80 mM or 150 mM NaCl after digestion contain predominantly mononucleosomes and represent classical "active" chromatin. Profiles of these low-salt soluble fractions display phased nucleosomes over transcriptionally active genes that are locally depleted of histone H3.3 and correspond closely to profiles of histone H2Av (H2A.Z) and RNA polymerase II. This correspondence suggests that transcription can result in loss of H3.3+H2Av nucleosomes and generate low-salt soluble nucleosomes. Nearly quantitative recovery of chromatin is obtained with 600 mM NaCl; however, the remaining insoluble chromatin is enriched in actively transcribed regions. Salt-insoluble chromatin likely represents oligonucleosomes that are attached to large protein complexes. Both low-salt extracted and insoluble chromatin are rich in sequences that correspond to epigenetic regulatory elements genome-wide. The presence of active chromatin at both extremes of salt solubility suggests that these salt fractions capture bound and unbound intermediates in active processes, thus providing a simple, powerful strategy for mapping epigenome dynamics.
Figures
References
-
- Ahmad K., Henikoff S. The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol. Cell. 2002;9:1191–1200. - PubMed
-
- Annunziato A.T., Schindler R.K., Thomas C.A., Jr, Seale R.L. Dual nature of newly replicated chromatin. Evidence for nucleosomal and non-nucleosomal DNA at the site of native replication forks. J. Biol. Chem. 1981;256:11880–11886. - PubMed
-
- Ausubel F.M., Brent R., Kingston R.E., Moore D.D., Seidman J.G., Smith J.A., Struhl K. Current protocols in molecular biology. Wiley; New York: 1994.
-
- Barski A., Cuddapah S., Cui K., Roh T.Y., Schones D.E., Wang Z., Wei G., Chepelev I., Zhao K. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129:823–837. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases