

Activator protein 2 alpha (AP2alpha) suppresses 42 kDa C/CAAT enhancer binding protein alpha (p42(C/EBPalpha)) in head and neck squamous cell carcinoma
- PMID: 19089912
- PMCID: PMC2676424
- DOI: 10.1002/ijc.24087
Activator protein 2 alpha (AP2alpha) suppresses 42 kDa C/CAAT enhancer binding protein alpha (p42(C/EBPalpha)) in head and neck squamous cell carcinoma
Abstract
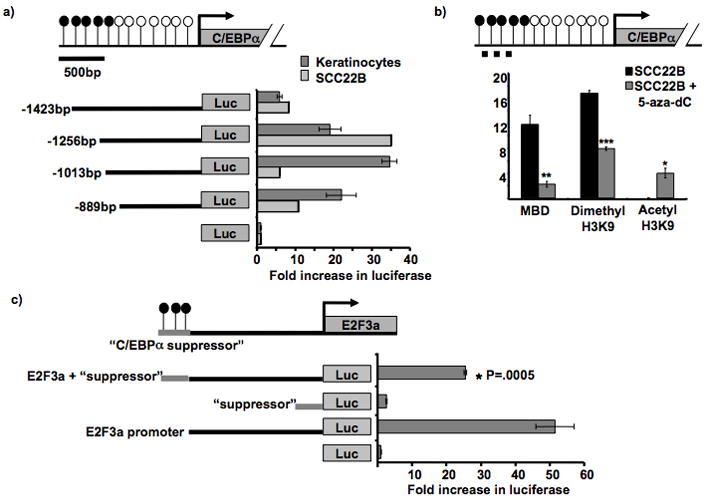
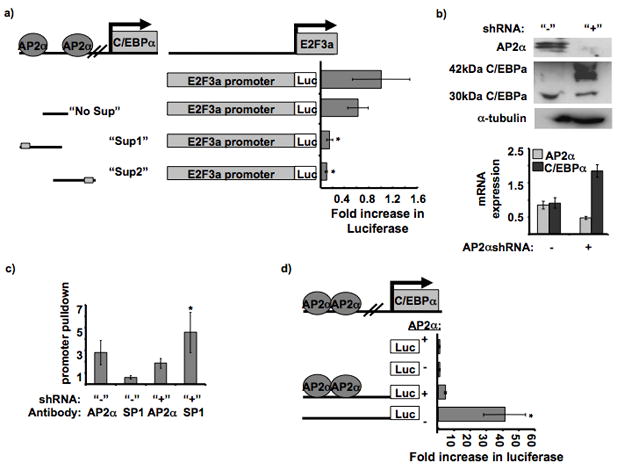
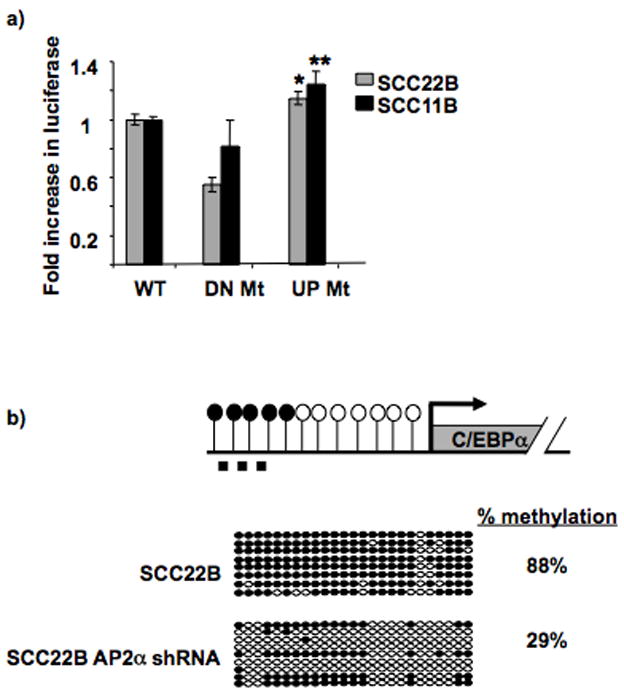
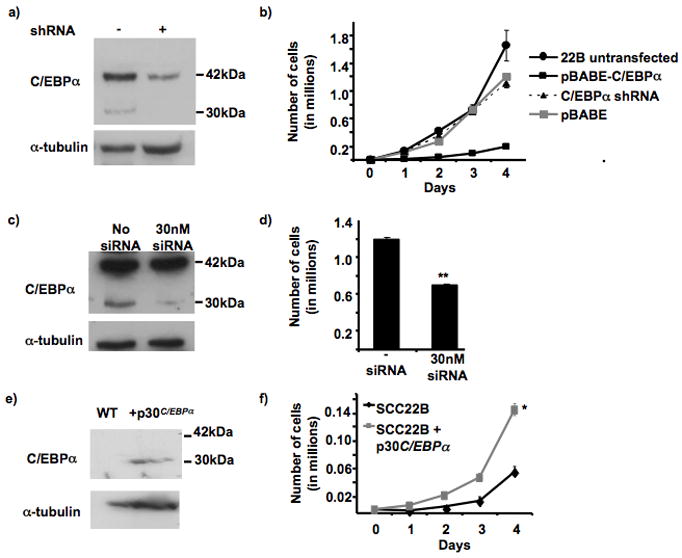
The tumor suppressor C/CAAT enhancer binding protein alpha (C/EBPalpha) is a transcription factor involved in cell cycle control and cellular differentiation. A recent study showed that C/EBPalpha is frequently downregulated in head and neck squamous cell carcinoma (HNSCC) by DNA methylation in an upstream regulatory region. Here, we investigated how DNA methylation in the upstream regulatory region disrupts the transcriptional regulation of C/EBPalpha in HNSCC. The results reveal that aberrant methylation correlates with methyl binding domain protein binding and repressive histone modifications. This methylated region contains previously uninvestigated AP2alpha binding sites. AP2alpha suppresses C/EBPalpha promoter activity and protein expression. Interestingly, silencing AP2alpha by shRNA increases the antiproliferative isoform of C/EBPalpha (p42(C/EBPalpha)). Furthermore, growth analysis revealed that these 2 isoforms yield very different proliferative properties in HNSCC.
Figures
References
-
- Schrem H, Klempnauer J, Borlak J. Liver-enriched transcription factors in liver function and development. Part II: the C/EBPs and D site-binding protein in cell cycle control, carcinogenesis, circadian gene regulation, liver regeneration, apoptosis, and liver-specific gene regulation. Pharmacol Rev. 2004;56:291–330. - PubMed
-
- Johnson PF. Molecular stop signs: regulation of cell-cycle arrest by C/EBP transcription factors. J Cell Sci. 2005;118:2545–55. - PubMed
-
- Hsieh CC, Xiong W, Xie Q, Rabek JP, Scott SG, An MR, Reisner PD, Kuninger DT, Papaconstantinou J. Effects of age on the posttranscriptional regulation of CCAAT/enhancer binding protein alpha and CCAAT/enhancer binding protein beta isoform synthesis in control and LPS-treated livers. Mol Biol Cell. 1998;9:1479–94. - PMC - PubMed
-
- Halmos B, Huettner CS, Kocher O, Ferenczi K, Karp DD, Tenen DG. Down-regulation and antiproliferative role of C/EBPalpha in lung cancer. Cancer Res. 2002;62:528–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
