

Mechanism of formation of the C-terminal beta-hairpin of the B3 domain of the immunoglobulin binding protein G from Streptococcus. I. Importance of hydrophobic interactions in stabilization of beta-hairpin structure
- PMID: 19089955
- PMCID: PMC2791351
- DOI: 10.1002/prot.22304
Mechanism of formation of the C-terminal beta-hairpin of the B3 domain of the immunoglobulin binding protein G from Streptococcus. I. Importance of hydrophobic interactions in stabilization of beta-hairpin structure
Abstract
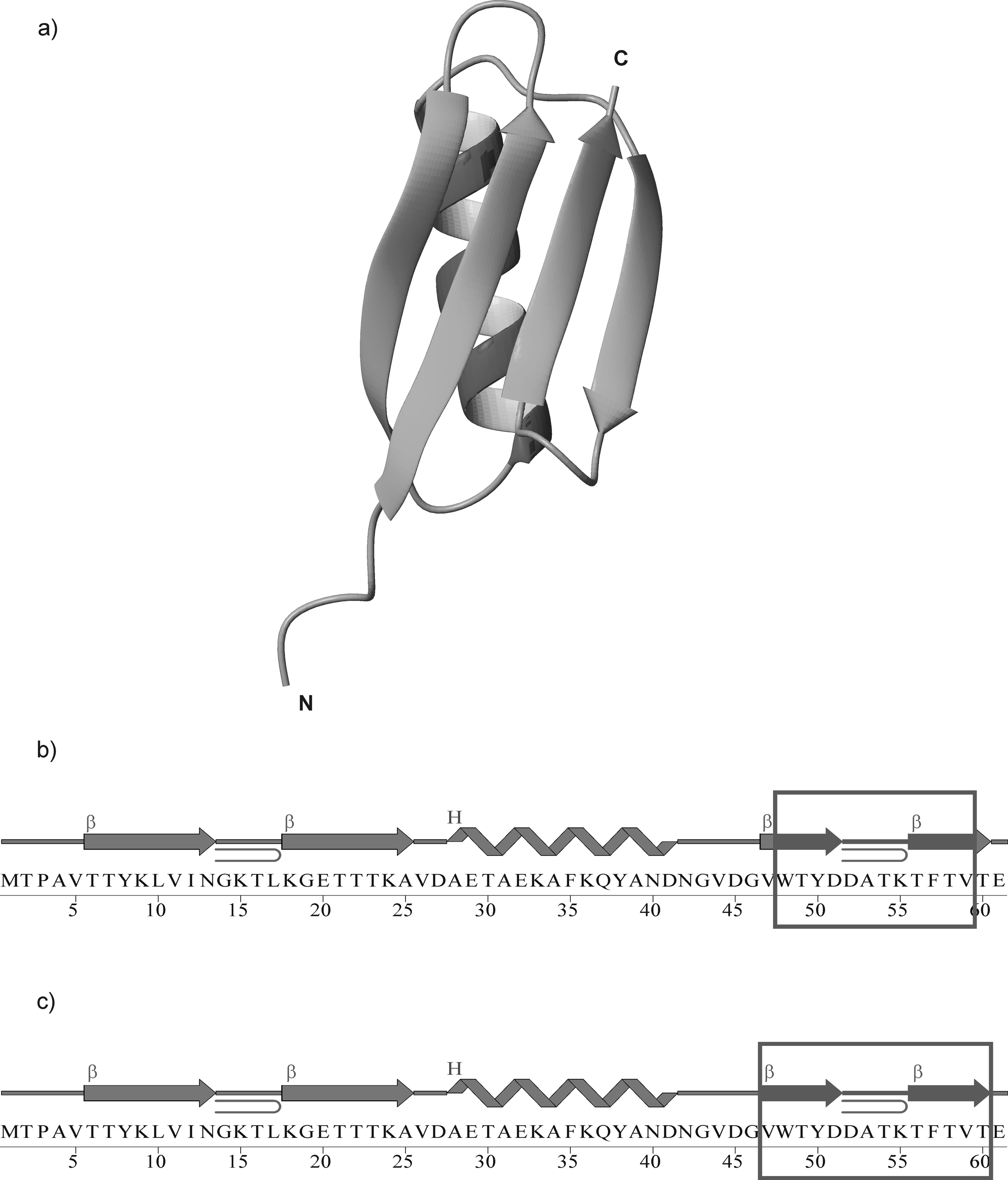
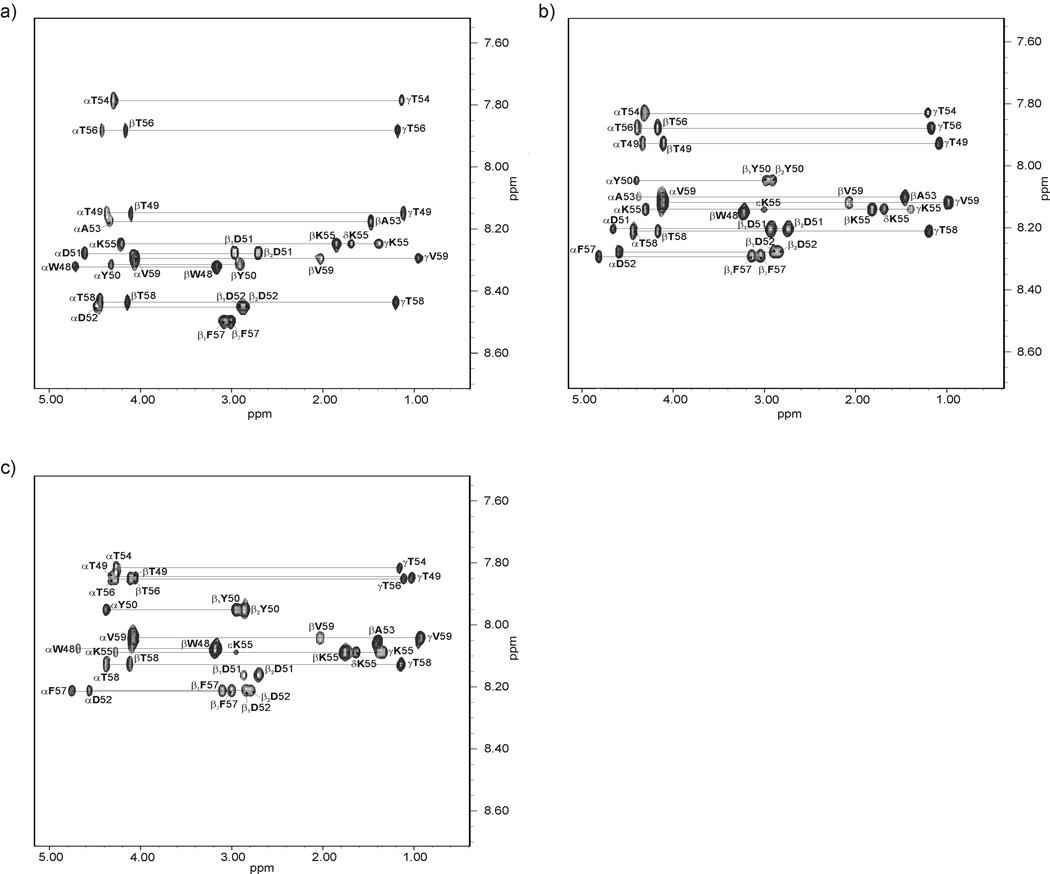
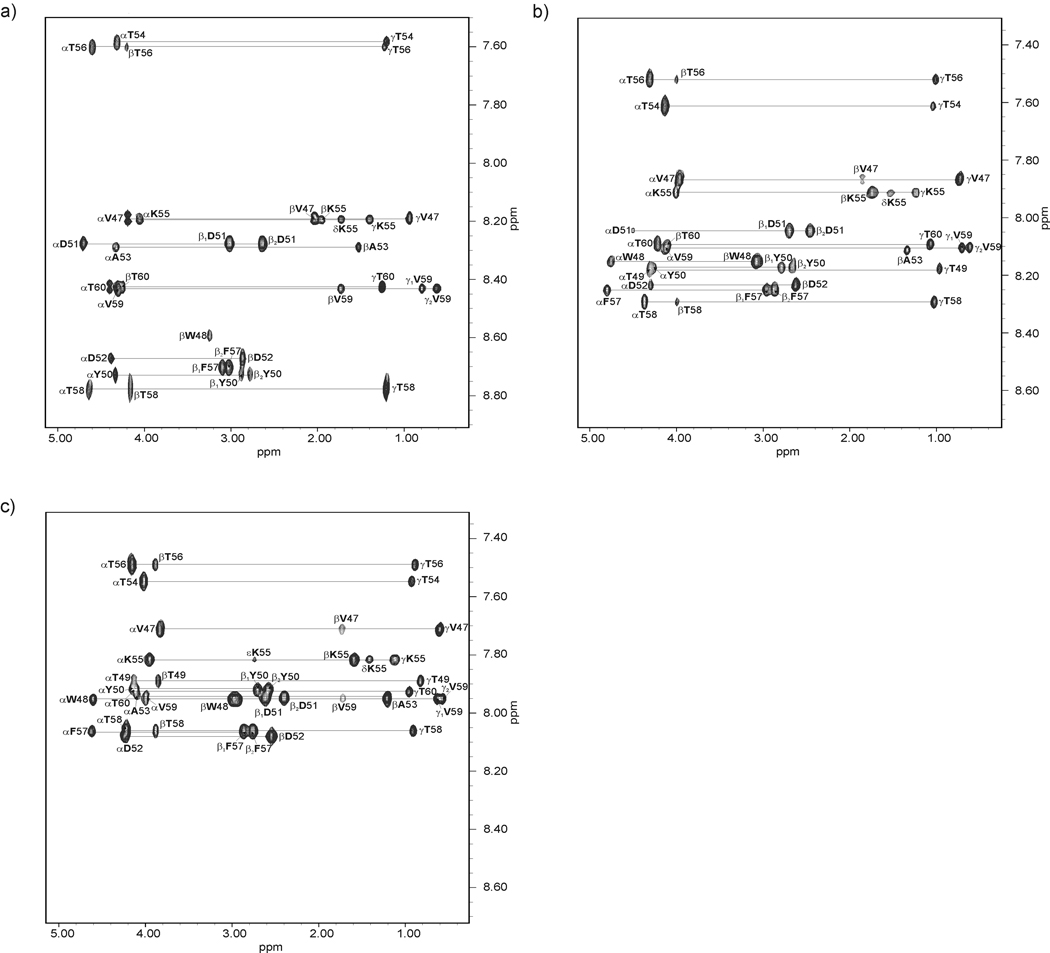
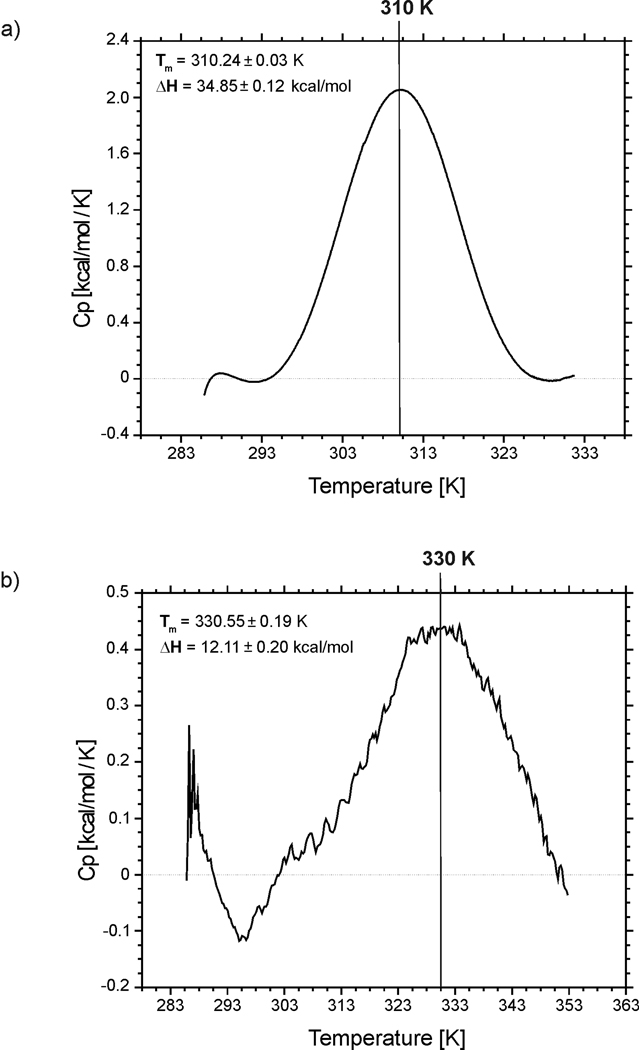
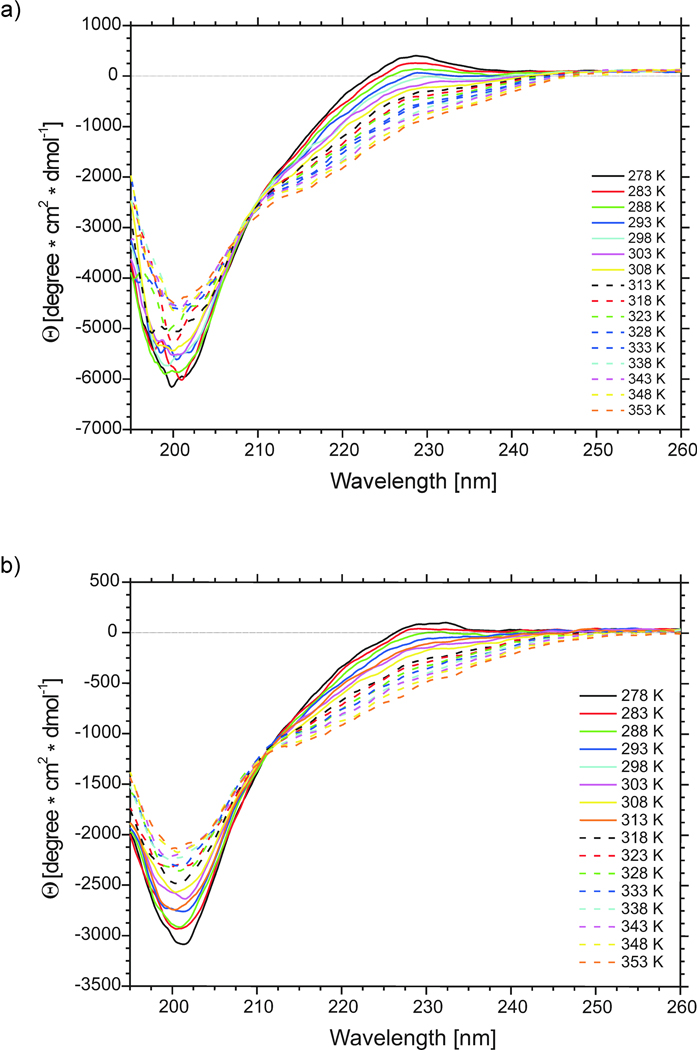
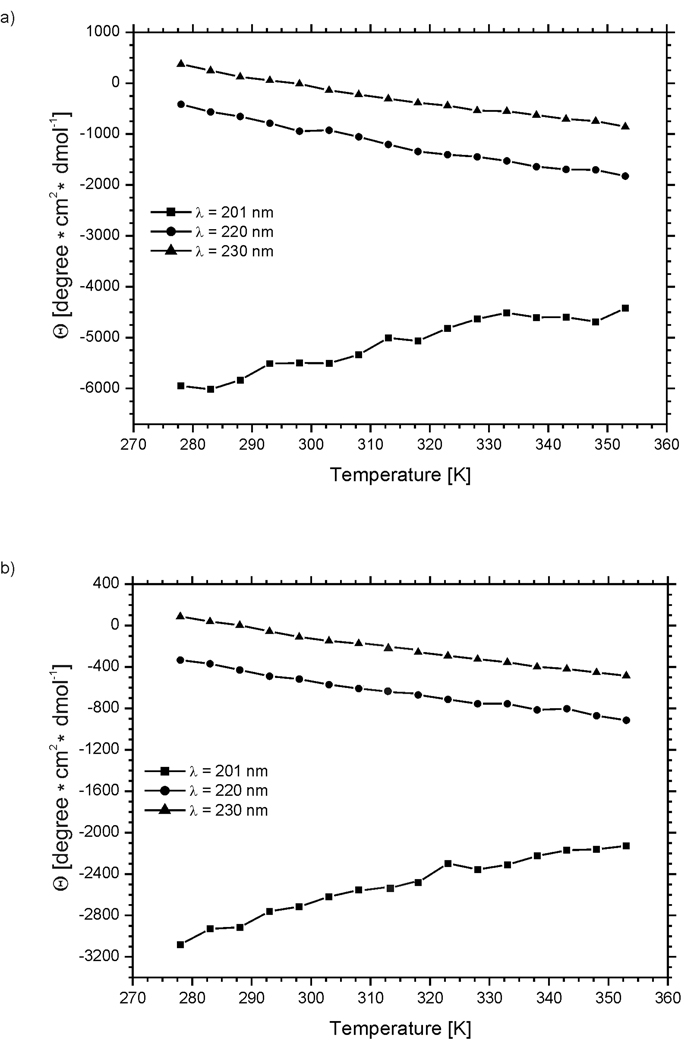
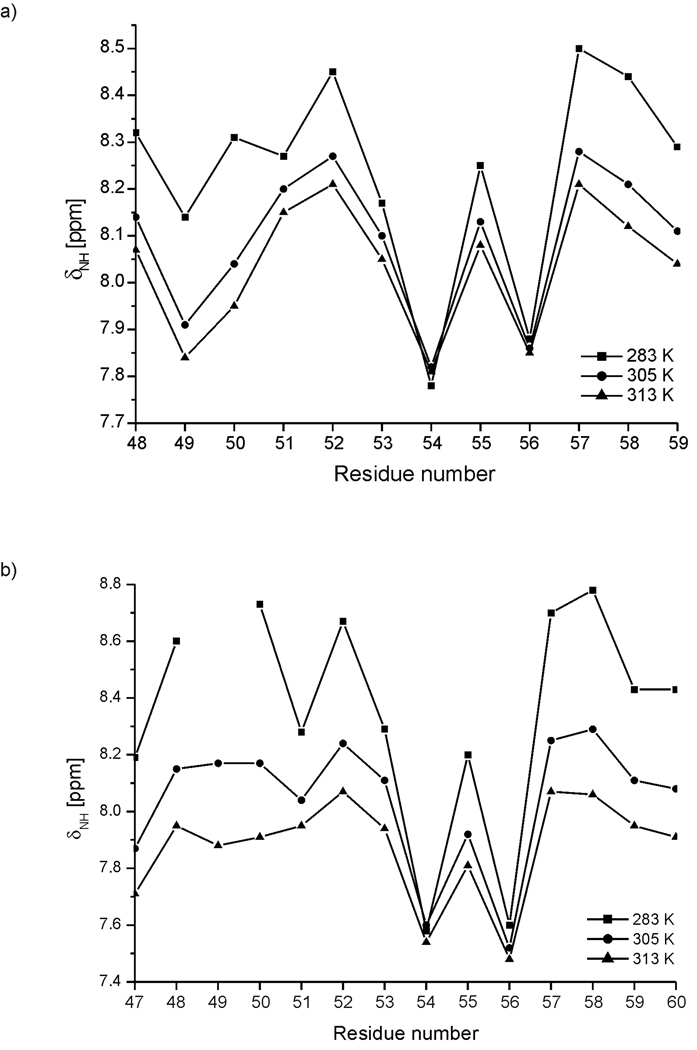
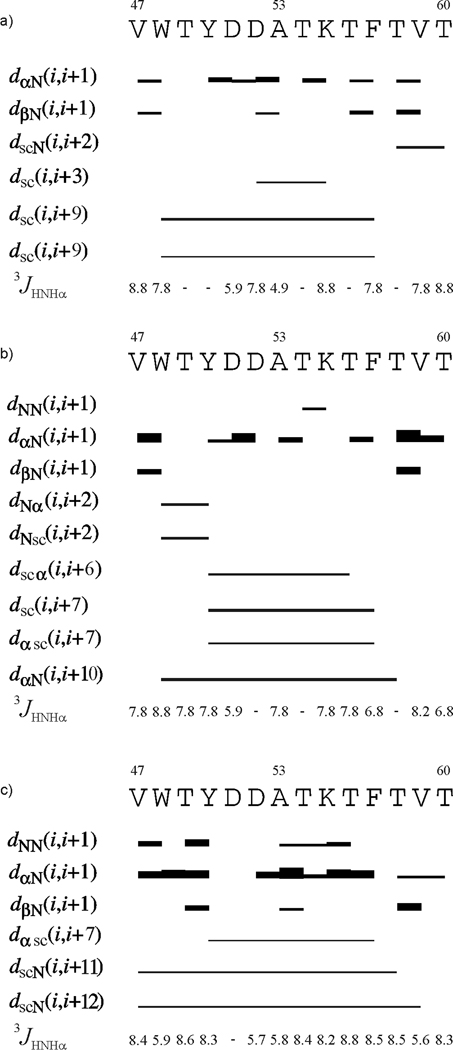
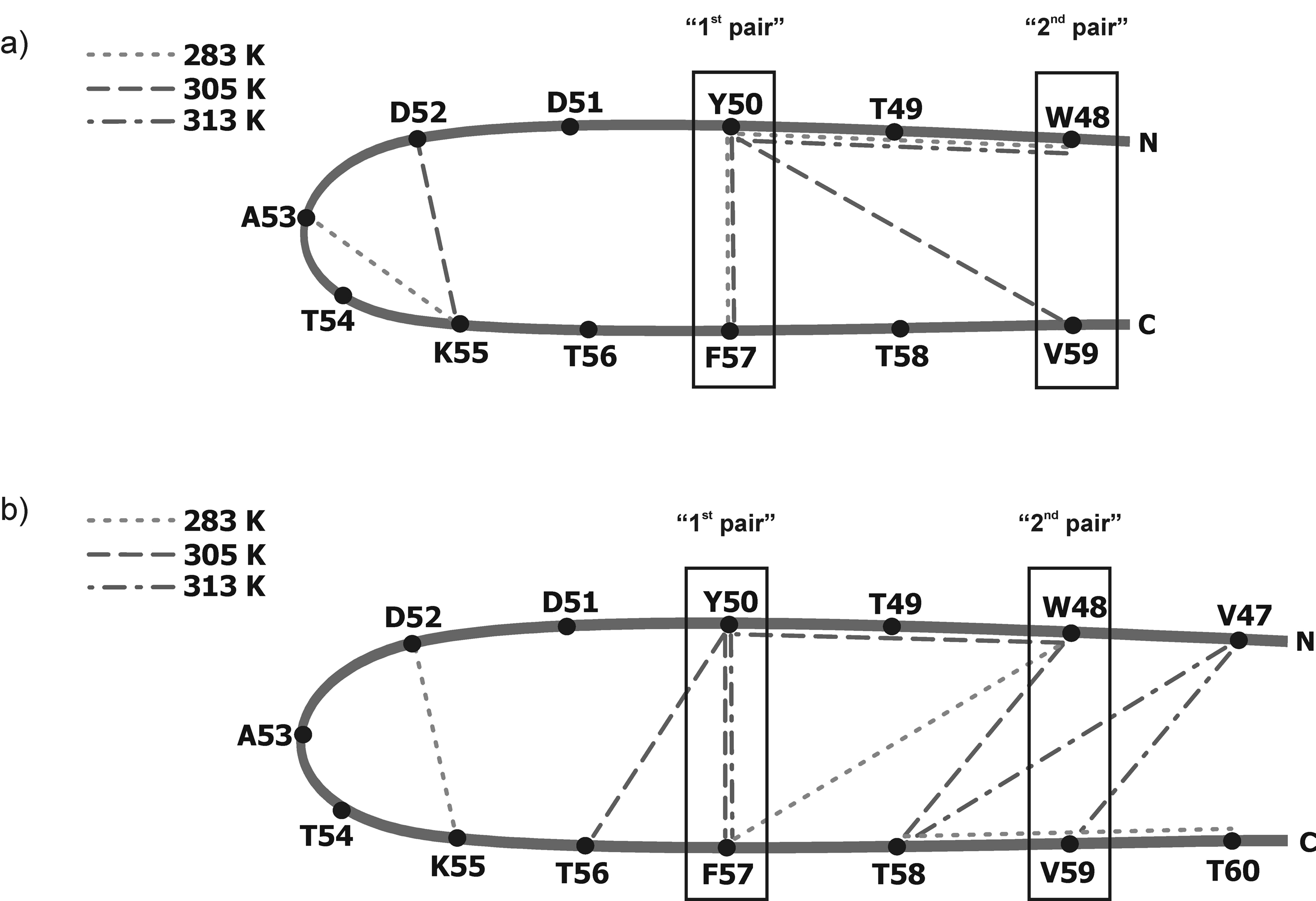
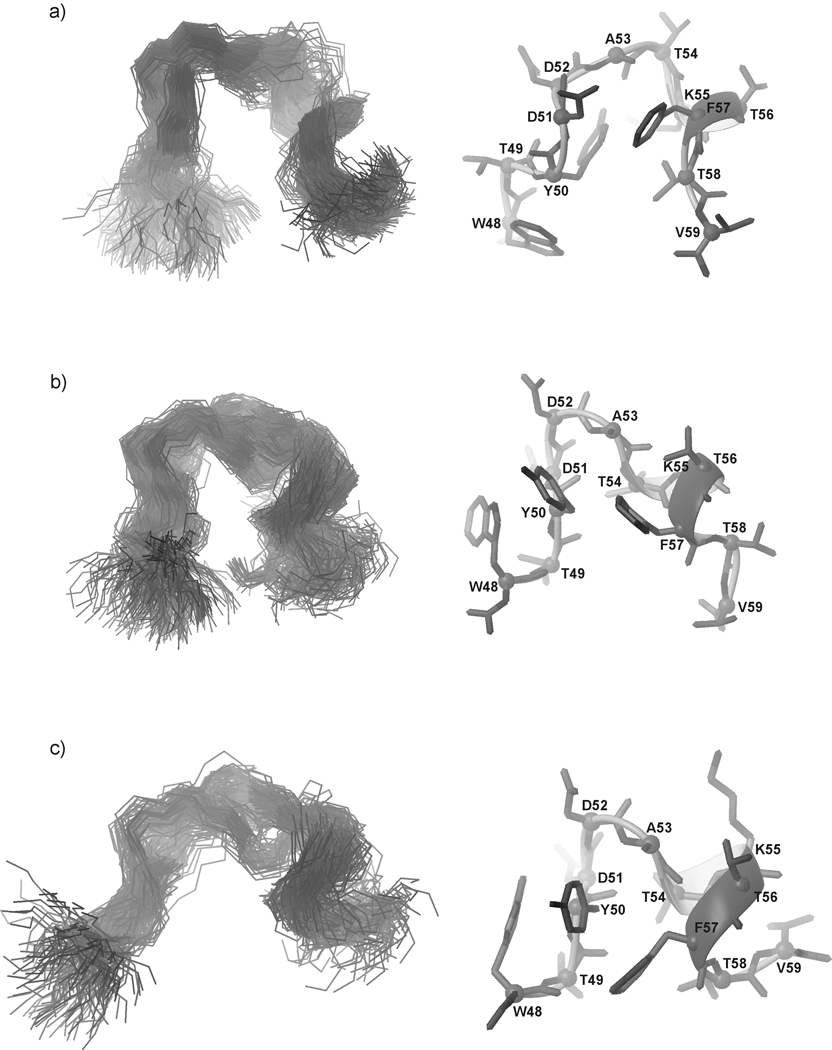
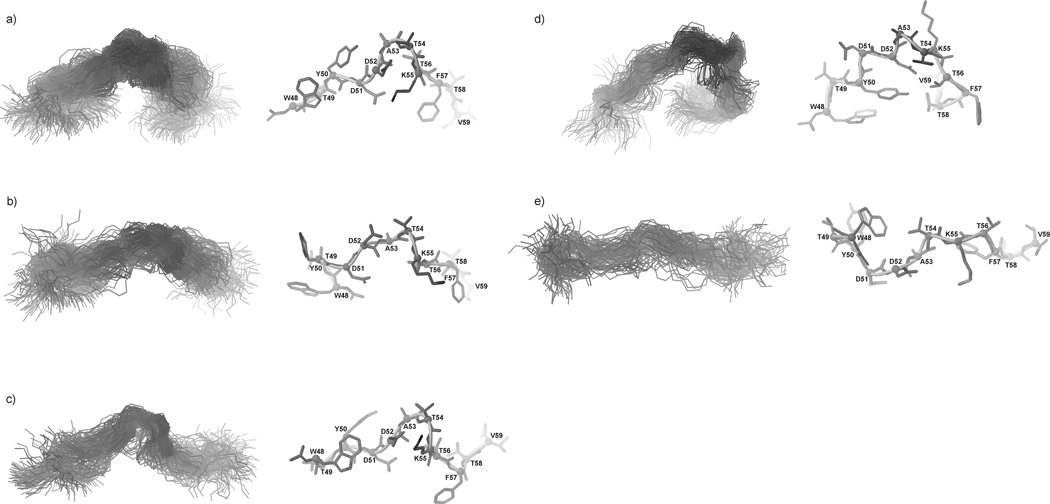
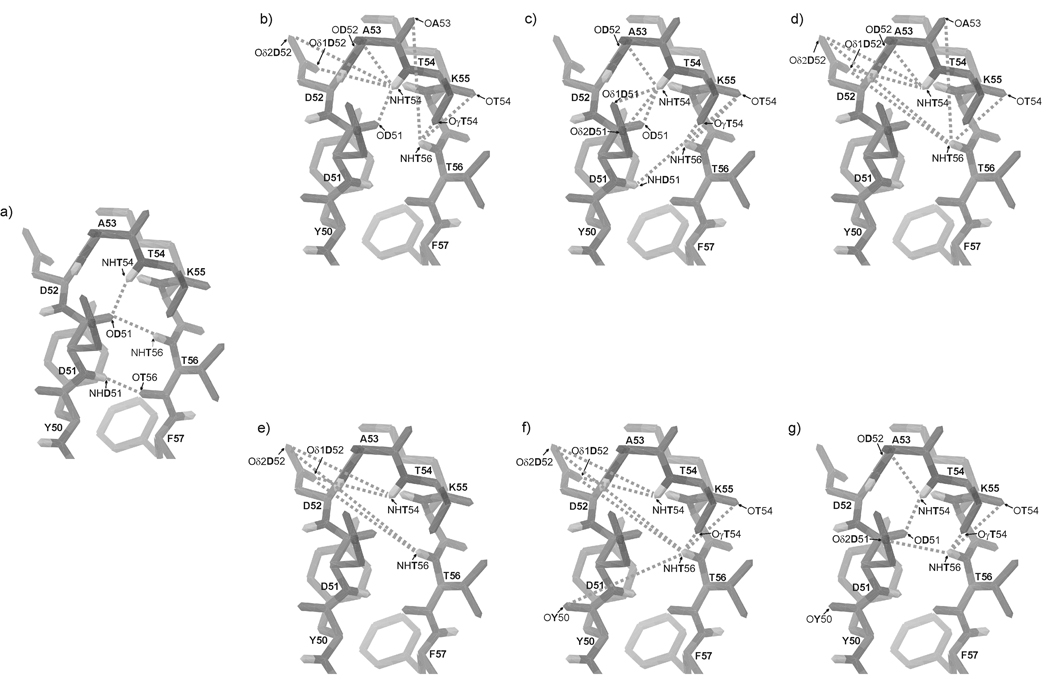
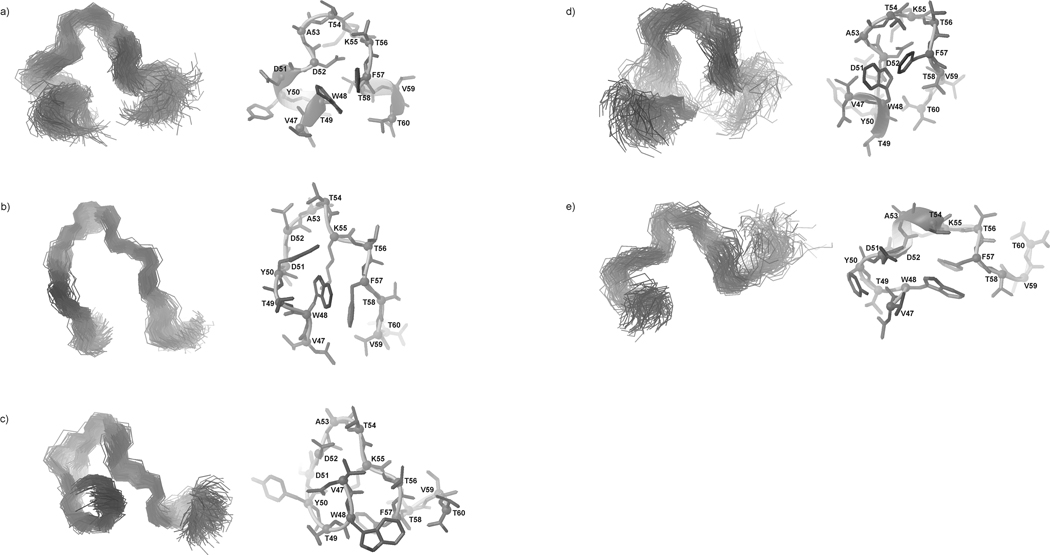
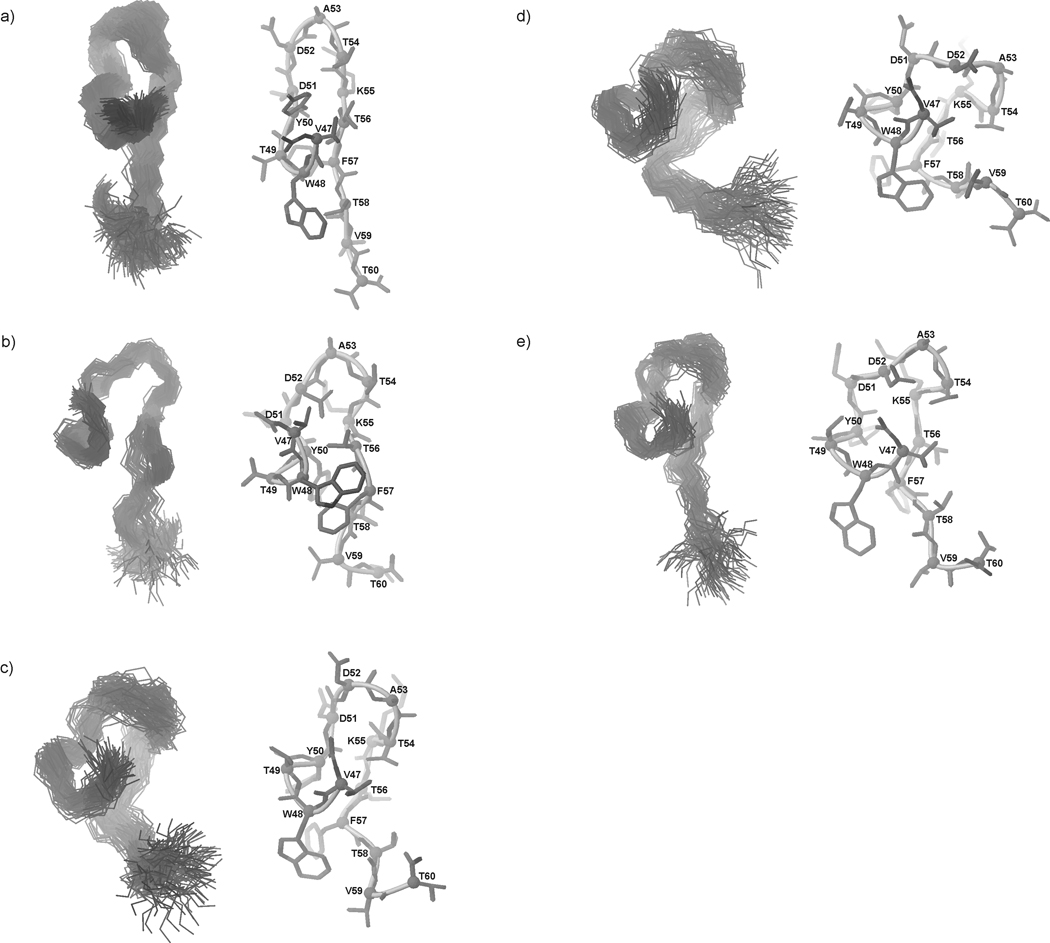
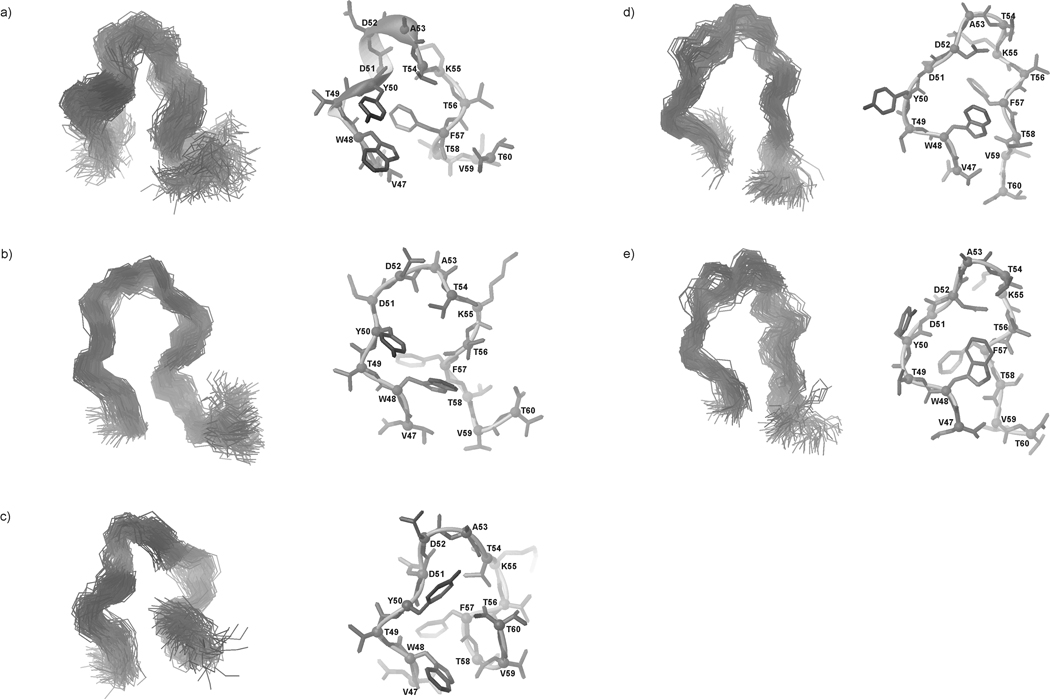
We previously studied a 16-amino acid-residue fragment of the C-terminal beta-hairpin of the B3 domain (residues 46-61), [IG(46-61)] of the immunoglobulin binding protein G from Streptoccocus, and found that hydrophobic interactions and the turn region play an important role in stabilizing the structure. Based on these results, we carried out systematic structural studies of peptides derived from the sequence of IG (46-61) by systematically shortening the peptide by one residue at a time from both the C- and the N-terminus. To determine the structure and stability of two resulting 12- and 14-amino acid-residue peptides, IG(48-59) and IG(47-60), respectively, we carried out circular dichroism, NMR, and calorimetric studies of these peptides in pure water. Our results show that IG(48-59) possesses organized three-dimensional structure stabilized by hydrophobic interactions (Tyr50-Phe57 and Trp48-Val59) at T = 283 and 305 K. At T = 313 K, the structure breaks down because of increased chain entropy, but the turn region is preserved in the same position observed for the structure of the whole protein. The breakdown of structure occurs near the melting temperature of this peptide (T(m) = 310 K) measured by differential scanning calorimetry (DSC). The melting temperature of IG(47-60) determined by DSC is T(m) = 330 K and its structure is similar to that of the native beta-hairpin at all (lower) temperatures examined (283-313 K). Both of these truncated sequences are conserved in all known amino acid sequences of the B domains of the immunoglobulin binding protein G from bacteria. Thus, this study contributes to an understanding of the mechanism of folding of this whole family of proteins, and provides information about the mechanism of formation and stabilization of a beta-hairpin structural element.
Copyright 2008 Wiley-Liss, Inc.
Figures

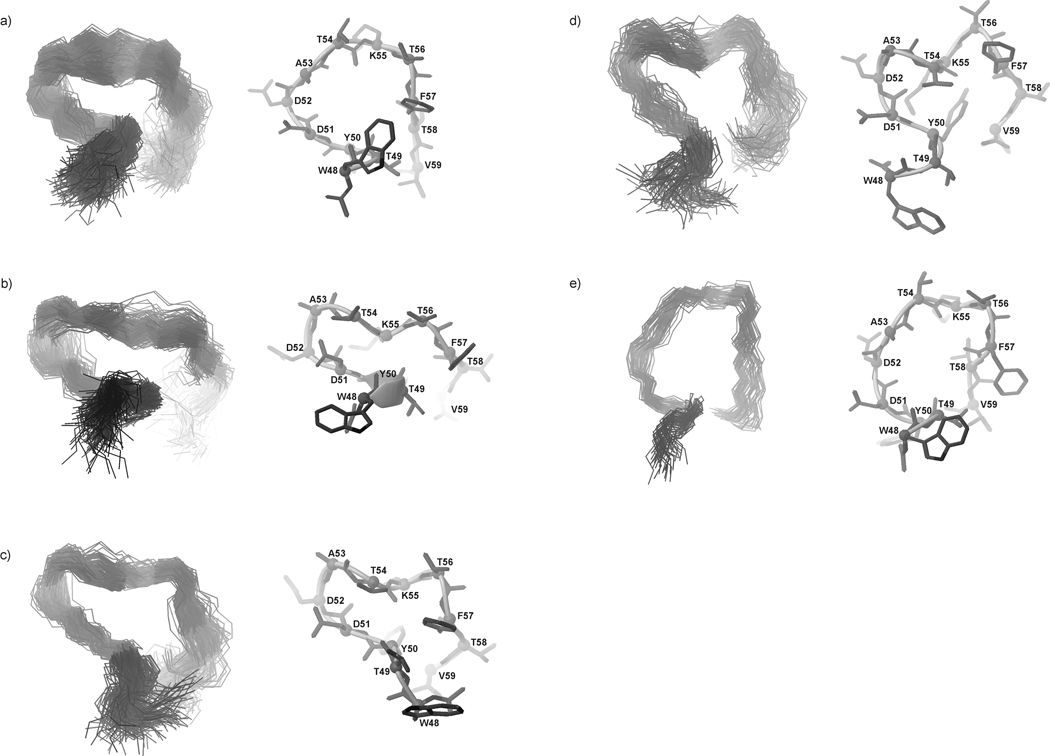
References
-
- Matheson RR, Jr, Scheraga HA. A method for predicting nucleation sites for protein folding based on hydrophobic contacts. Macromolecules. 1978;11:819–829.
-
- Matheson RR, Jr, Scheraga HA. Steps in the pathway of the thermal unfolding of ribonuclease A. A nonspecific surface-labeling study. Biochemistry. 1979;12:2437–2445. - PubMed
-
- Honig B. Protein folding: From the Levinthal paradox to structure prediction. J Mol Biol. 1999;293:283–293. - PubMed
-
- Baldwin RL, Rose GD. Is protein folding hierarchic? I. Local structure and peptide folding. Trends Biochem Sci. 1999;24:26–33. - PubMed
-
- Baldwin RL, Rose GD. Is protein folding hierarchic? II. Folding intermediates and transition states. Trends Biochem Sci. 1999;24:77–83. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
