

Root system architecture from coupling cell shape to auxin transport
- PMID: 19090618
- PMCID: PMC2602721
- DOI: 10.1371/journal.pbio.0060307
Root system architecture from coupling cell shape to auxin transport
Erratum in
- PLoS Biol. 2014 Oct;12(10):e1001984
Abstract
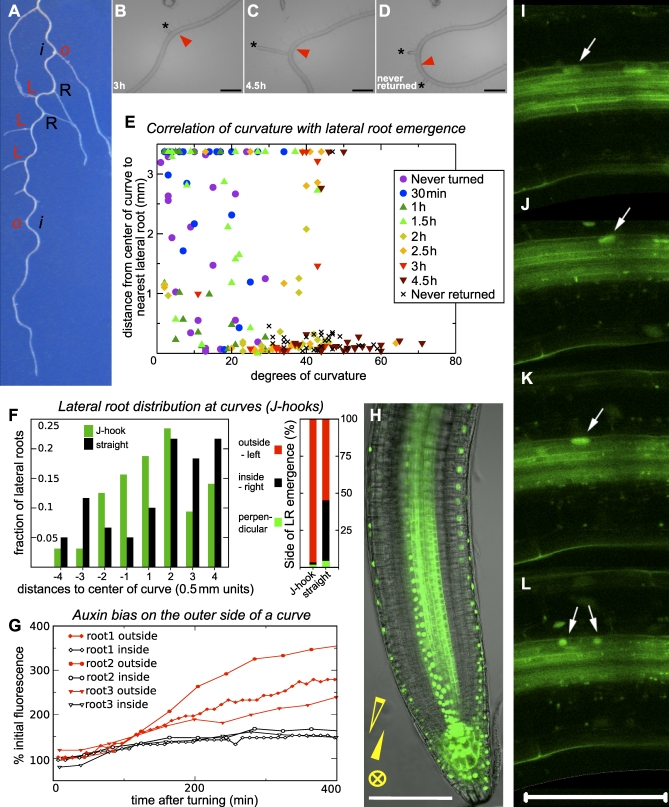
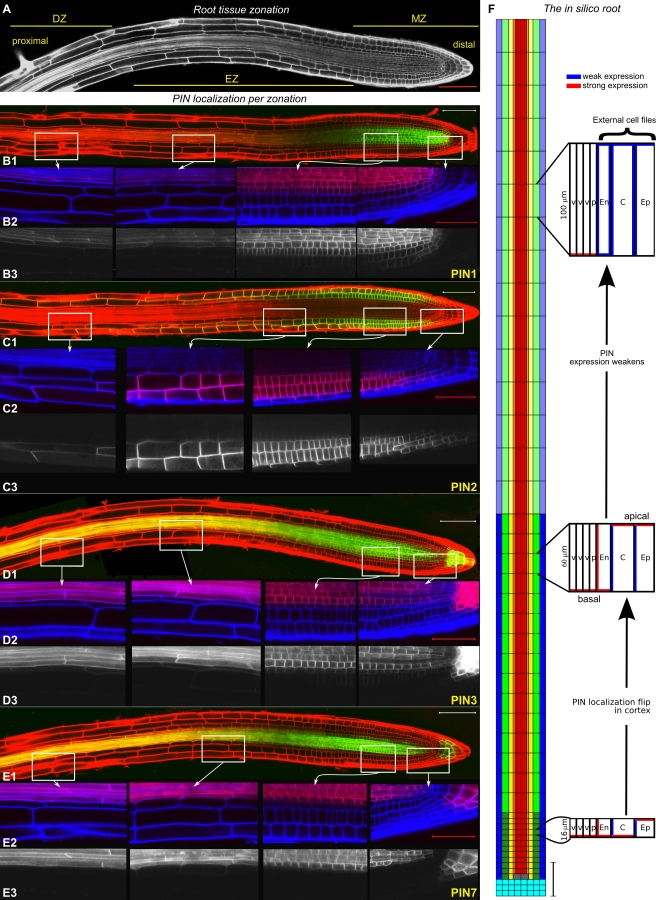
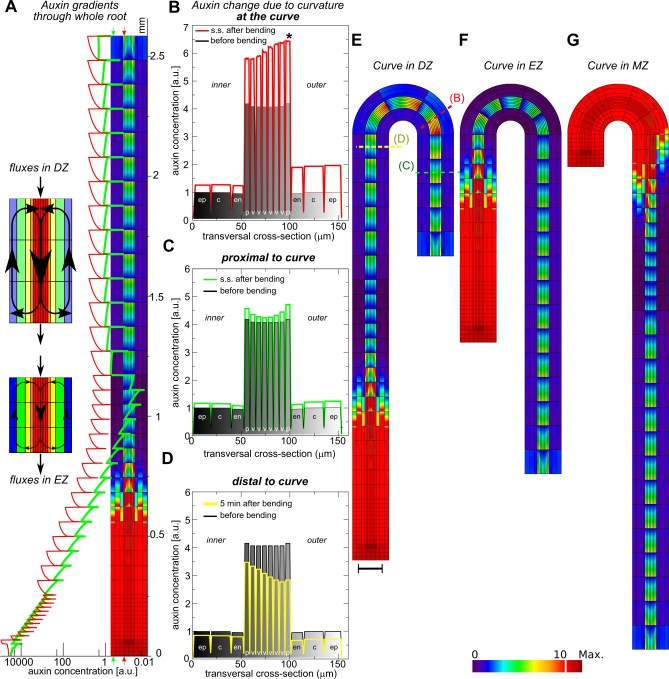
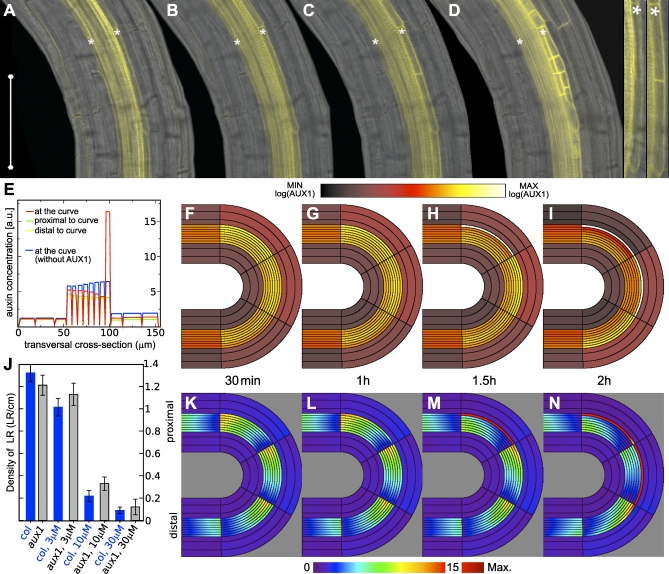
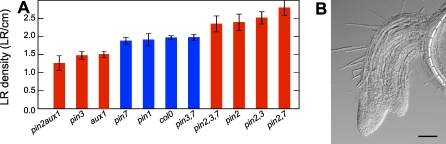
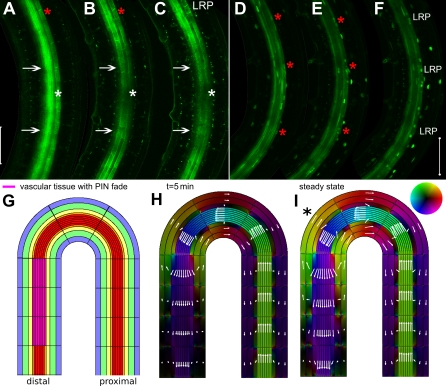
Lateral organ position along roots and shoots largely determines plant architecture, and depends on auxin distribution patterns. Determination of the underlying patterning mechanisms has hitherto been complicated because they operate during growth and division. Here, we show by experiments and computational modeling that curvature of the Arabidopsis root influences cell sizes, which, together with tissue properties that determine auxin transport, induces higher auxin levels in the pericycle cells on the outside of the curve. The abundance and position of the auxin transporters restricts this response to the zone competent for lateral root formation. The auxin import facilitator, AUX1, is up-regulated by auxin, resulting in additional local auxin import, thus creating a new auxin maximum that triggers organ formation. Longitudinal spacing of lateral roots is modulated by PIN proteins that promote auxin efflux, and pin2,3,7 triple mutants show impaired lateral inhibition. Thus, lateral root patterning combines a trigger, such as cell size difference due to bending, with a self-organizing system that mediates alterations in auxin transport.
Conflict of interest statement
Figures
Comment in
-
The role of auxin transport in plant patterning mechanisms.PLoS Biol. 2008 Dec 16;6(12):e323. doi: 10.1371/journal.pbio.0060323. PLoS Biol. 2008. PMID: 19090623 Free PMC article.
References
-
- Van Tieghem P, Duliot H. Recherches comparatives sur l'origine des membres endogènes dans les plantes vasculaires. Ann Sci Nat Bot. 1888;7:1–660.
-
- Fortin MC, Pierce FJ, Poff KL. The pattern of secondary root formation in curving roots of Arabidopsis thaliana (L.) Heynh. Plant Cell Environ. 1989;12:337–339. - PubMed
-
- De Smet I, Tetsumura T, De Rybel B, Frei dit Frey N, Laplaze L, et al. Auxin-dependent regulation of lateral root positioning in the basal meristem of Arabidopsis . Development. 2007;134:681–690. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
