

Prox1 maintains muscle structure and growth in the developing heart
- PMID: 19091769
- PMCID: PMC2655234
- DOI: 10.1242/dev.030007
Prox1 maintains muscle structure and growth in the developing heart
Erratum in
- Development. 2009 Feb;136(4):699
Abstract
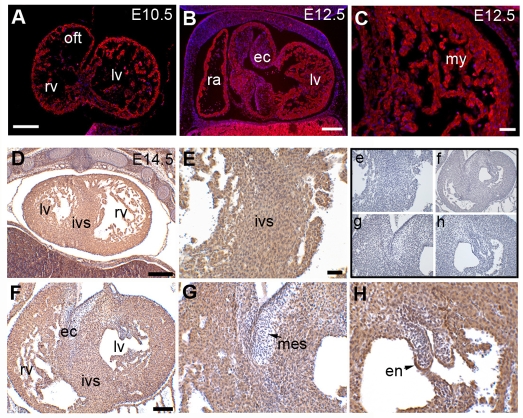
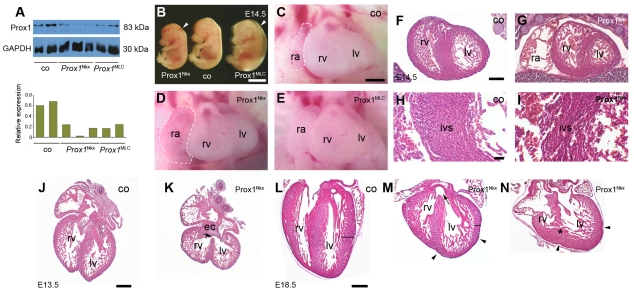
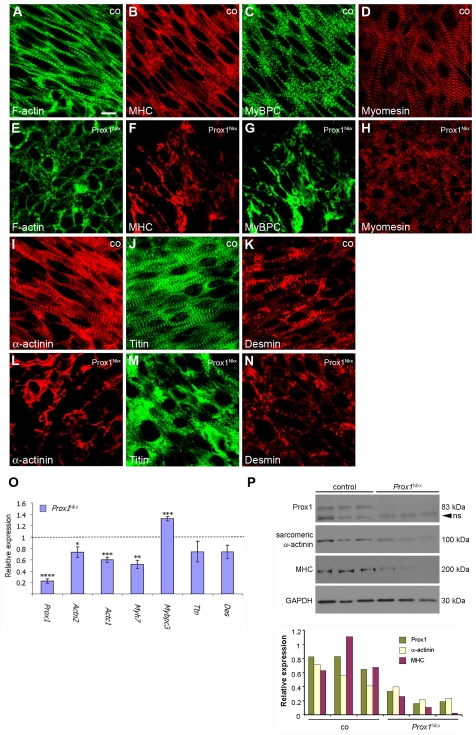
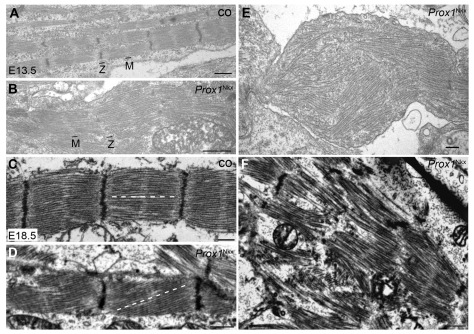
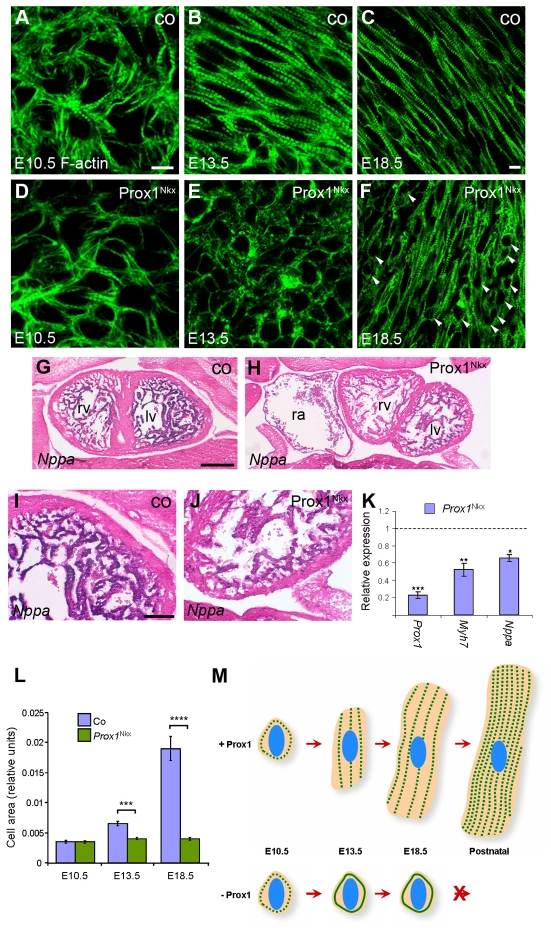
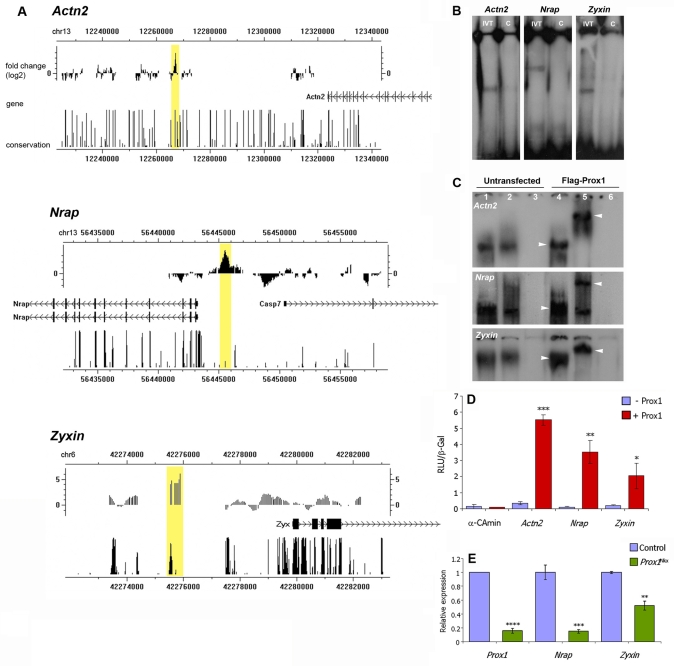
Impaired cardiac muscle growth and aberrant myocyte arrangement underlie congenital heart disease and cardiomyopathy. We show that cardiac-specific inactivation of the murine homeobox transcription factor Prox1 results in the disruption of expression and localisation of sarcomeric proteins, gross myofibril disarray and growth-retarded hearts. Furthermore, we demonstrate that Prox1 is required for direct transcriptional regulation of the genes encoding the structural proteins alpha-actinin, N-RAP and zyxin, which collectively function to maintain an actin-alpha-actinin interaction as the fundamental association of the sarcomere. Aspects of abnormal heart development and the manifestation of a subset of muscular-based disease have previously been attributed to mutations in key structural proteins. Our study reveals an essential requirement for direct transcriptional regulation of sarcomere integrity, in the context of enabling foetal cardiomyocyte hypertrophy, maintenance of contractile function and progression towards inherited or acquired myopathic disease.
Figures
References
-
- Ahuja, P., Perriard, E., Perriard, J. C. and Ehler, E. (2004). Sequential myofibrillar breakdown accompanies mitotic division of mammalian cardiomyocytes. J. Cell Sci. 117, 3295-3306. - PubMed
-
- Akazawa, H. and Komuro, I. (2003). Roles of cardiac transcription factors in cardiac hypertrophy. Circ. Res. 92, 1079-1088. - PubMed
-
- Alcalai, R., Seidman, J. G. and Seidman, C. E. (2008). Genetic basis of hypertrophic cardiomyopathy: from bench to the clinics. J. Cardiovasc. Electrophysiol. 19, 104-110. - PubMed
-
- Angelo, S., Lohr, J., Lee, K. H., Ticho, B. S., Breitbart, R. E., Hill, S., Yost, H. J. and Srivastava, D. (2000). Conservation of sequence and expression of Xenopus and zebrafish dHAND during cardiac, branchial arch and lateral mesoderm development. Mech. Dev. 95, 231-237. - PubMed
-
- Balza, R. O., Jr and Misra, R. P. (2006). Role of the serum response factor in regulating contractile apparatus gene expression and sarcomeric integrity in cardiomyocytes. J. Biol. Chem. 281, 6498-6510. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
