

Efferent-mediated responses in vestibular nerve afferents of the alert macaque
- PMID: 19091917
- PMCID: PMC2657077
- DOI: 10.1152/jn.91112.2008
Efferent-mediated responses in vestibular nerve afferents of the alert macaque
Abstract
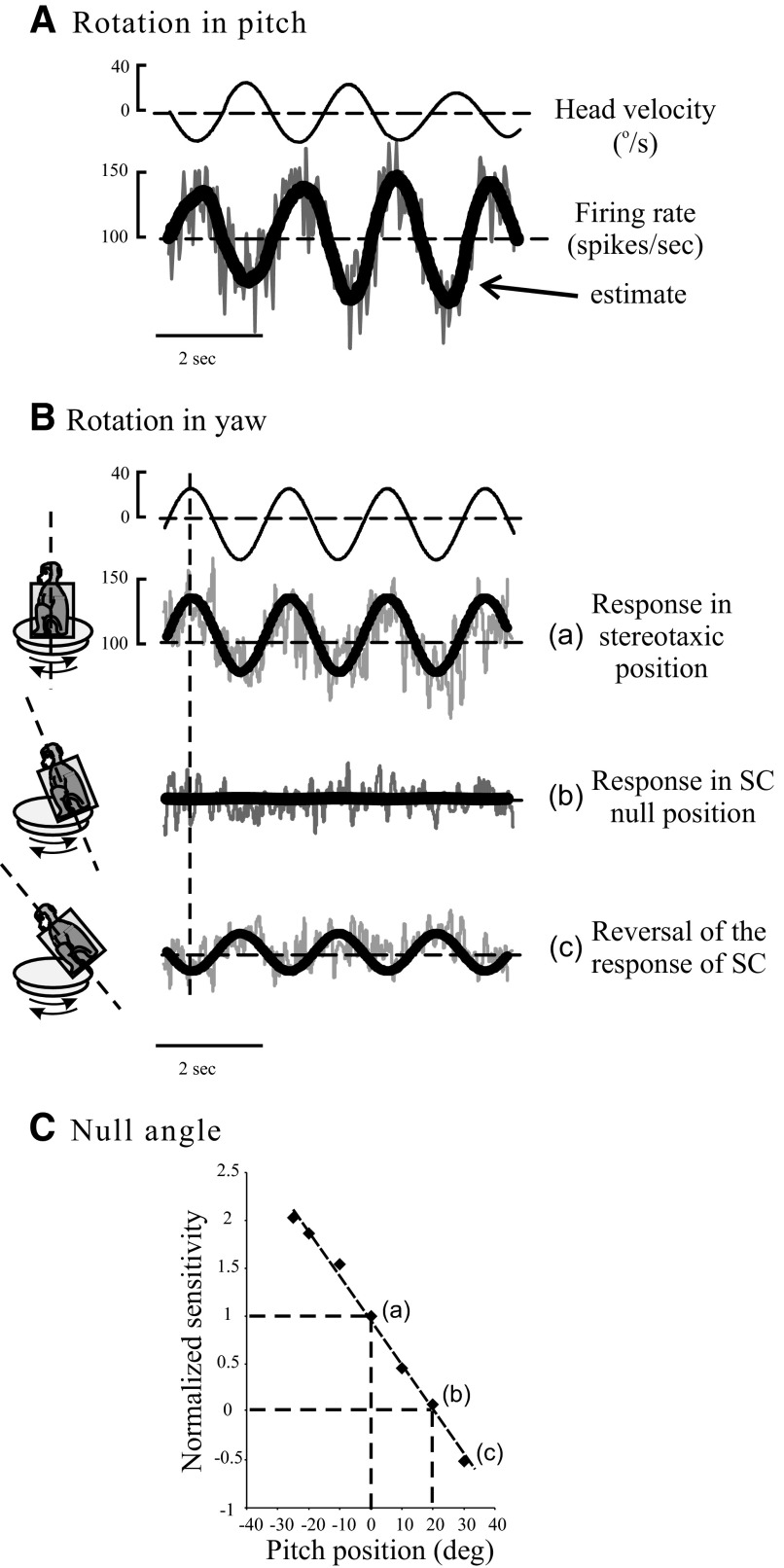
The peripheral vestibular organs have long been known to receive a bilateral efferent innervation from the brain stem. However, the functional role of the efferent vestibular system has remained elusive. In this study, we investigated efferent-mediated responses in vestibular afferents of alert behaving primates (macaque monkey). We found that efferent-mediated rotational responses could be obtained from vestibular nerve fibers innervating the semicircular canals after conventional afferent responses were nulled by placing the corresponding canal plane orthogonal to the plane of motion. Responses were type III, i.e., excitatory for rotational velocity trapezoids (peak velocity, 320 degrees/s) in both directions of rotation, consistent with those previously reported in the decerebrate chinchilla. Responses consisted of both fast and slow components and were larger in irregular (approximately 10 spikes/s) than in regular afferents (approximately 2 spikes/s). Following unilateral labyrinthectomy (UL) on the side opposite the recording site, similar responses were obtained. To confirm the vestibular source of the efferent-mediated responses, the ipsilateral horizontal and posterior canals were plugged following the UL. Responses to high-velocity rotations were drastically reduced when the superior canal (SC), the only intact canal, was in its null position, compared with when the SC was pitched 50 degrees upward from the null position. Our findings show that vestibular afferents in alert primates show efferent-mediated responses that are related to the discharge regularity of the afferent, are of vestibular origin, and can be the result of both afferent excitation and inhibition.
Figures






Similar articles
-
Reflections of efferent activity in rotational responses of chinchilla vestibular afferents.J Neurophysiol. 2002 Sep;88(3):1234-44. doi: 10.1152/jn.2002.88.3.1234. J Neurophysiol. 2002. PMID: 12205144
-
Semicircular canal afferents similarly encode active and passive head-on-body rotations: implications for the role of vestibular efference.J Neurosci. 2002 Jun 1;22(11):RC226. doi: 10.1523/JNEUROSCI.22-11-j0002.2002. Epub 2002 May 23. J Neurosci. 2002. PMID: 12040085 Free PMC article.
-
Response of vestibular-nerve afferents to active and passive rotations under normal conditions and after unilateral labyrinthectomy.J Neurophysiol. 2007 Feb;97(2):1503-14. doi: 10.1152/jn.00829.2006. Epub 2006 Nov 22. J Neurophysiol. 2007. PMID: 17122313
-
Effects of canal plugging on the vestibuloocular reflex and vestibular nerve discharge during passive and active head rotations.J Neurophysiol. 2009 Nov;102(5):2693-703. doi: 10.1152/jn.00710.2009. Epub 2009 Sep 2. J Neurophysiol. 2009. PMID: 19726724 Free PMC article.
-
Determinants of spatial and temporal coding by semicircular canal afferents.J Neurophysiol. 2005 May;93(5):2359-70. doi: 10.1152/jn.00533.2004. J Neurophysiol. 2005. PMID: 15845995 Free PMC article. Review.
Cited by
-
Specializations for Fast Signaling in the Amniote Vestibular Inner Ear.Integr Comp Biol. 2018 Aug 1;58(2):341-350. doi: 10.1093/icb/icy069. Integr Comp Biol. 2018. PMID: 29920589 Free PMC article. Review.
-
Reviewing the Role of the Efferent Vestibular System in Motor and Vestibular Circuits.Front Physiol. 2017 Aug 2;8:552. doi: 10.3389/fphys.2017.00552. eCollection 2017. Front Physiol. 2017. PMID: 28824449 Free PMC article. Review.
-
Loss of α-Calcitonin Gene-Related Peptide (αCGRP) Reduces Otolith Activation Timing Dynamics and Impairs Balance.Front Mol Neurosci. 2018 Aug 24;11:289. doi: 10.3389/fnmol.2018.00289. eCollection 2018. Front Mol Neurosci. 2018. PMID: 30197585 Free PMC article.
-
Efferent synaptic transmission at the vestibular type II hair cell synapse.J Neurophysiol. 2020 Aug 1;124(2):360-374. doi: 10.1152/jn.00143.2020. Epub 2020 Jul 1. J Neurophysiol. 2020. PMID: 32609559 Free PMC article.
-
Implantable Direct Current Neural Modulation: Theory, Feasibility, and Efficacy.Front Neurosci. 2019 Apr 18;13:379. doi: 10.3389/fnins.2019.00379. eCollection 2019. Front Neurosci. 2019. PMID: 31057361 Free PMC article. Review.
References
-
- Baird RA, Desmadryl G, Fernandez C, Goldberg JM. The vestibular nerve of the chinchilla. II. Relation between afferent response properties and peripheral innervation patterns in the semicircular canals. J Neurophysiol 60: 182–203, 1988. - PubMed
-
- Bernard C, Cochran SL, Precht W. Presynaptic actions of cholinergic agents upon the hair cell-afferent fiber synapse in the vestibular labyrinth of the frog. Brain Res 338: 225–236, 1985. - PubMed
-
- Blanks RH, Precht W. Functional characterization of primary vestibular afferents in the frog. Exp Brain Res 25: 369–390, 1976. - PubMed
-
- Boyle R, Carey JP, Highstein SM. Morphological correlates of response dynamics and efferent stimulation in horizontal semicircular canal afferents of the toadfish, Opsanus tau. J Neurophysiol 66: 1504–1521, 1991. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources