

Adrenal peripheral clock controls the autonomous circadian rhythm of glucocorticoid by causing rhythmic steroid production
- PMID: 19091946
- PMCID: PMC2634940
- DOI: 10.1073/pnas.0806962106
Adrenal peripheral clock controls the autonomous circadian rhythm of glucocorticoid by causing rhythmic steroid production
Abstract
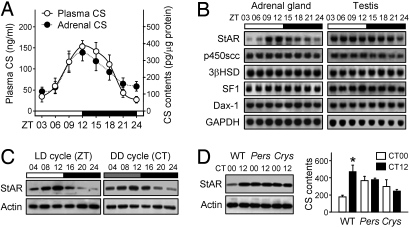
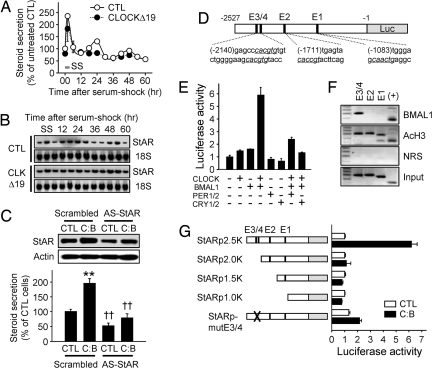
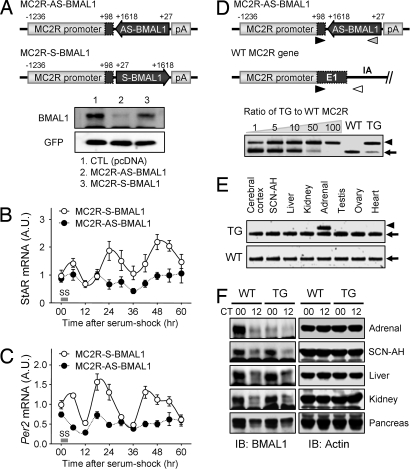
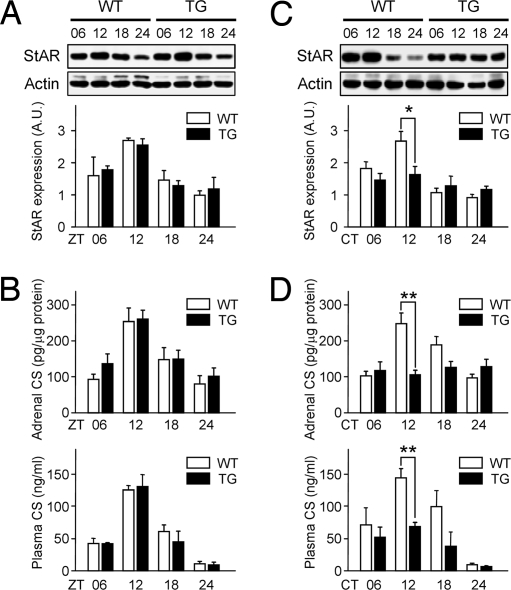
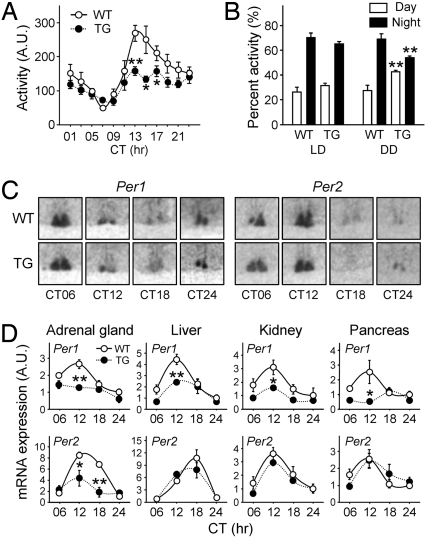
Glucocorticoid (GC) is an adrenal steroid with diverse physiological effects. It undergoes a robust daily oscillation, which has been thought to be driven by the master circadian clock in the suprachiasmatic nucleus of the hypothalamus via the hypothalamus-pituitary-adrenal axis. However, we show that the adrenal gland has its own clock and that the peripheral clockwork is tightly linked to steroidogenesis by the steroidogenic acute regulatory protein. Examination of mice with adrenal-specific knockdown of the canonical clock protein BMAL1 reveals that the adrenal clock machinery is required for circadian GC production. Furthermore, behavioral rhythmicity is drastically affected in these animals, together with altered expression of Period1, but not Period2, in several peripheral organs. We conclude that the adrenal peripheral clock plays an essential role in harmonizing the mammalian circadian timing system by generating a robust circadian GC rhythm.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ko CH, Takahashi JS. Molecular components of the mammalian circadian clock. Hum Mol Genet. 2006;15:R271–R277. - PubMed
-
- Stratmann M, Schibler U. Properties, entrainment, and physiological functions of mammalian peripheral oscillators. J Biol Rhythms. 2006;21:494–506. - PubMed
-
- Balsalobre A, et al. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science. 2000;289:2344–2347. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
