

The novel epididymis-specific beta-galactosidase-like gene Glb1l4 is essential in epididymal development and sperm maturation in rats
- PMID: 19092116
- PMCID: PMC11027174
- DOI: 10.1095/biolreprod.108.071589
The novel epididymis-specific beta-galactosidase-like gene Glb1l4 is essential in epididymal development and sperm maturation in rats
Abstract
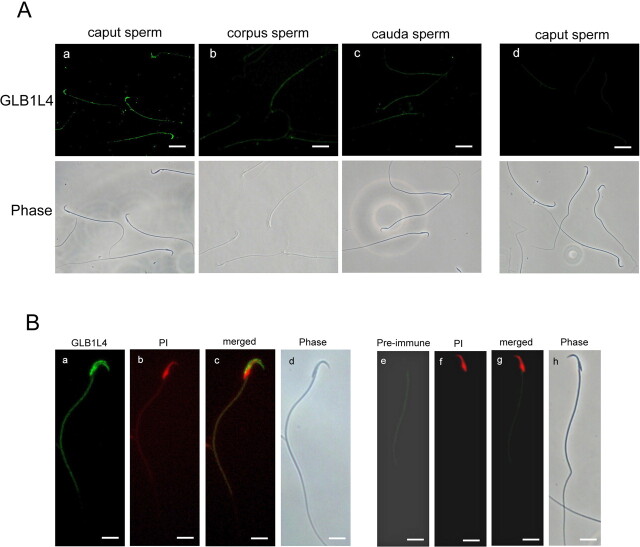
We describe a novel epididymis-specific cDNA named Glb1l4, which was isolated from rat epididymis by differential display of mRNAs. Glb1l4 cDNA contains 2607 nucleotides and encodes a 637-amino acid protein with 50% similarity to mouse beta-galactosidase. The gene is located on chromosome 8q13, spanning 21 exons. Northern blot analysis reveals that Glb1l4 is specifically expressed in the caput region of epididymis and upregulated by androgen. A specific polyclonal antiserum against the N-terminal peptide of GLB1L4 has been produced. Western blot analysis and immunohistochemistry assay reveal that GLB1L4 is specifically expressed in the principal cells of the caput epididymis. Interestingly, its expression peaks at Postnatal Day 45 in mRNA level and at Postnatal Day 60 in protein level while the epididymis column cells undergo differentiation. Moreover, within this very period this secretory protein is confined inside the cell with a change of subcellular distribution pattern, which implies its important roles in the cell differentiation process. Only after the epididymal epithelium differentiation is completed and the spermatozoa enter the epididymal lumen is the GLB1L4 secreted into the luminal fluid and bound on the sperm head. Our results suggest that GLB1L4 may play various roles in principal cell differentiation and sperm maturation.
Figures
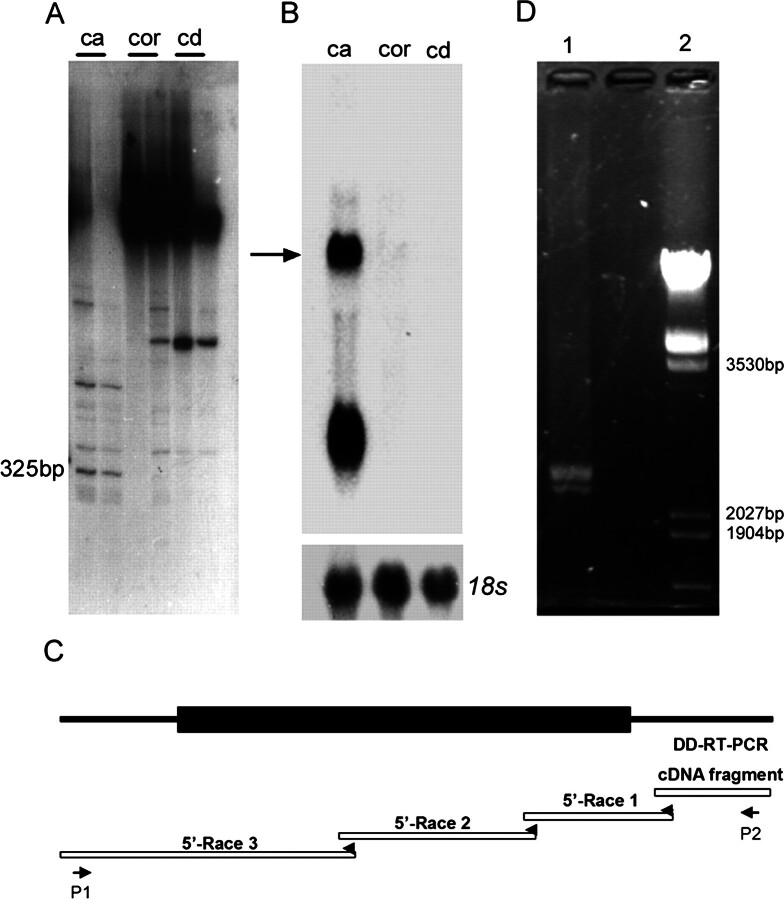


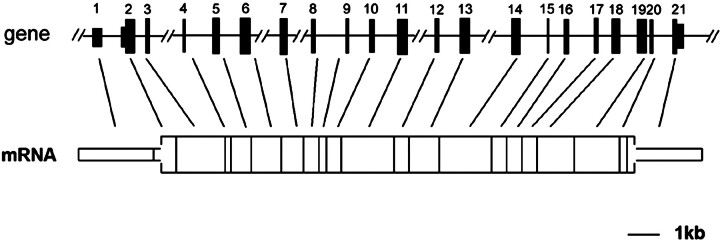
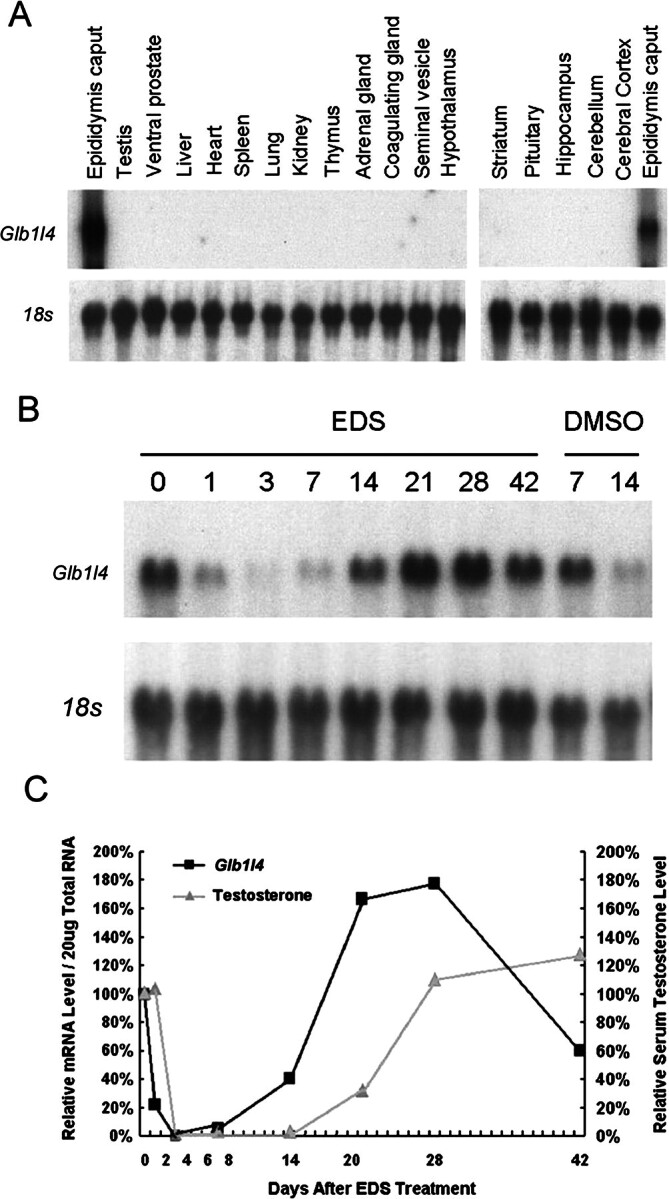
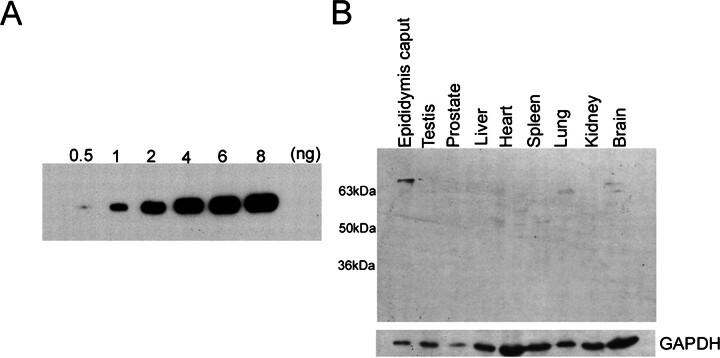
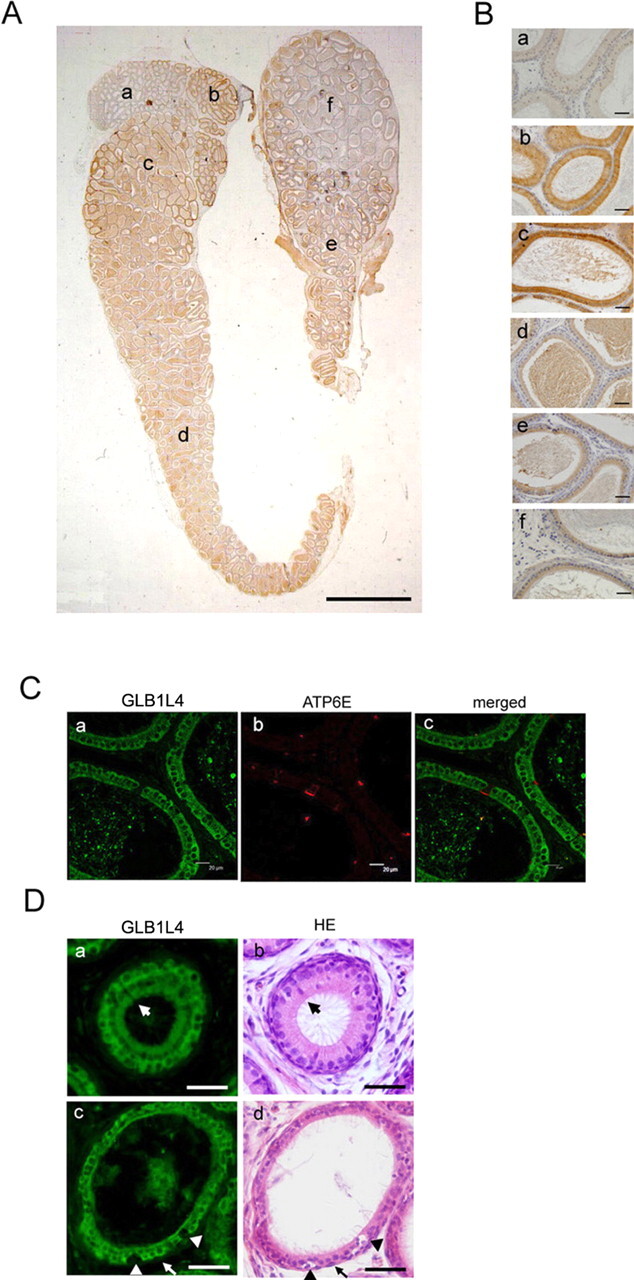
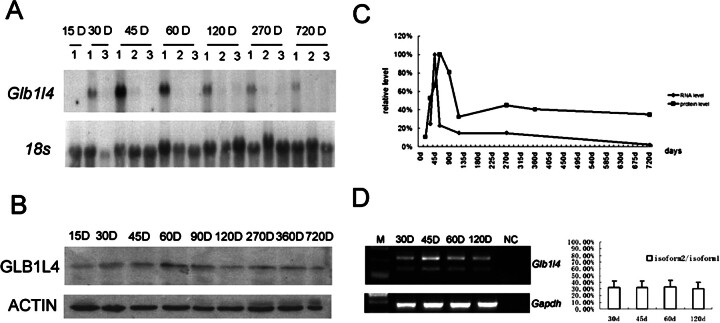
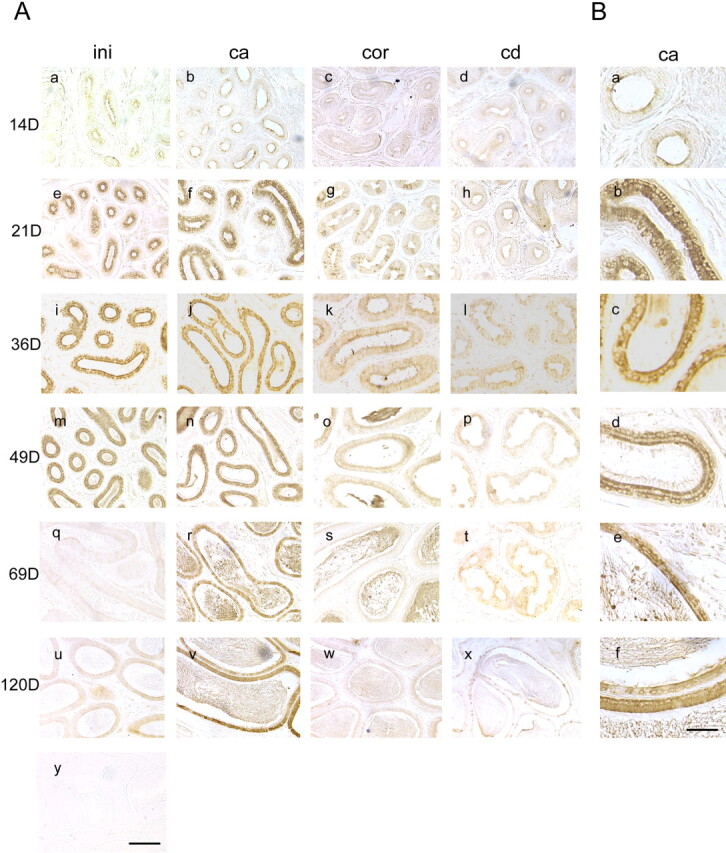
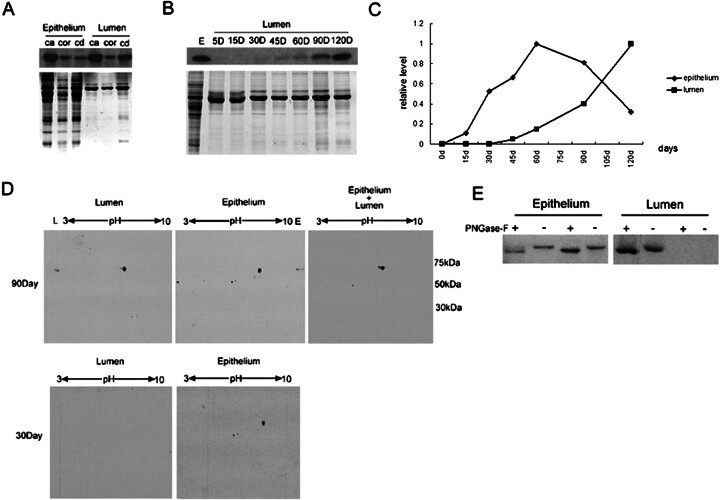
References
-
- Rodriguez CM, Kirby JL, Hinton BT. The development of the epididymis.In:Robaire B, Hinton BT. The Epididymis: From Molecules to Clinical Practice. New York: Kluwer Academic/Plenum Publishers; 2002:251–267.
-
- Connwall GA, Hann SR. Specialized gene expression in the epididymis. J Androl 1995; 16:379–383. - PubMed
-
- Hamilton DW. Structure and function of the epithelium lining the ductuli efferents, ducts epididymis and ductus deferens in the rat.In:Greep RO, Astwood EB. Handbook of Physiology, vol. 5. Baltimore: Waverly Press; 1975:259–383.
-
- Miler RJ, Killian GJ. Morphometric analysis of the epididymis from normal and vasectomized rats. J Androl 1987; 8:279–291. - PubMed
-
- Wong PYD, Au CL, Ngai HK. Electrolyte and water transport in rat epididymis: its possible role in sperm maturation. Int J Androl 1978; 1:600–628.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
