

The SV40 capsid is stabilized by a conserved pentapeptide hinge of the major capsid protein VP1
- PMID: 19094992
- PMCID: PMC2718546
- DOI: 10.1016/j.jmb.2008.11.034
The SV40 capsid is stabilized by a conserved pentapeptide hinge of the major capsid protein VP1
Abstract
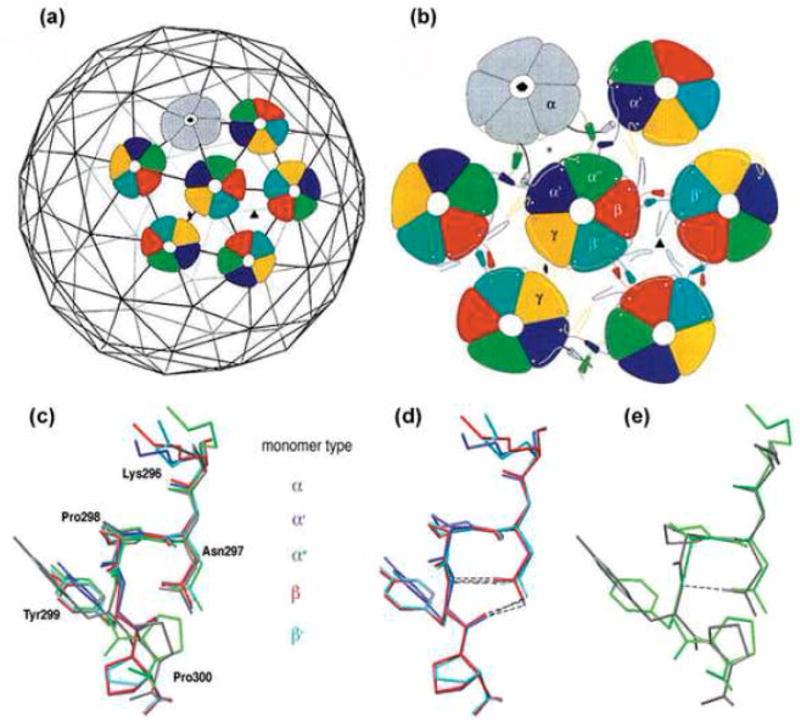
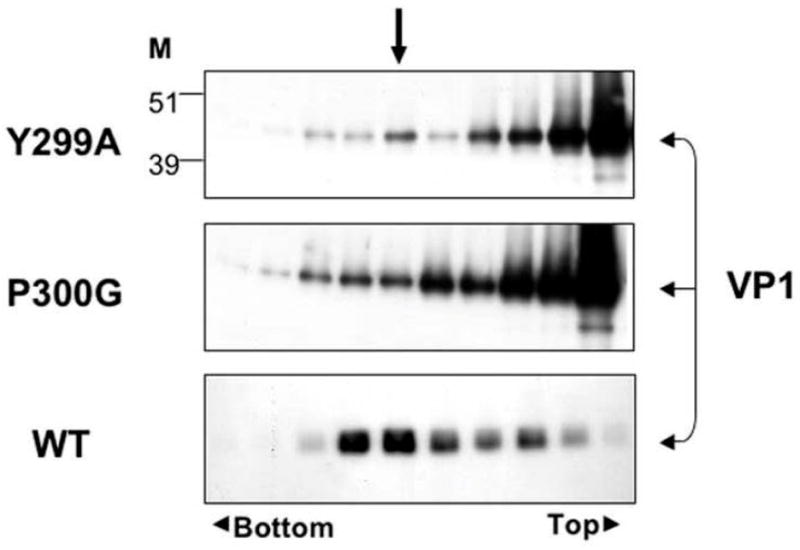
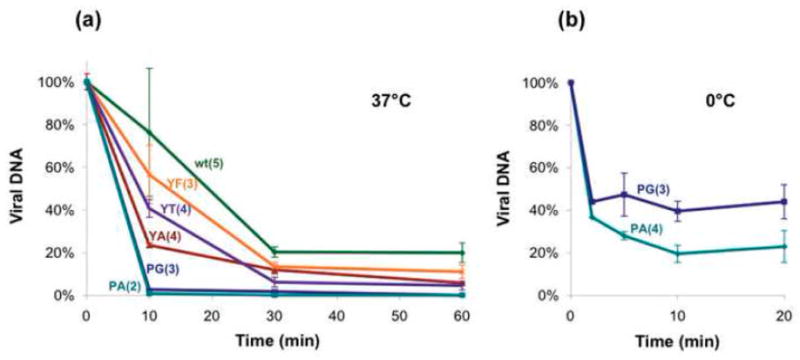
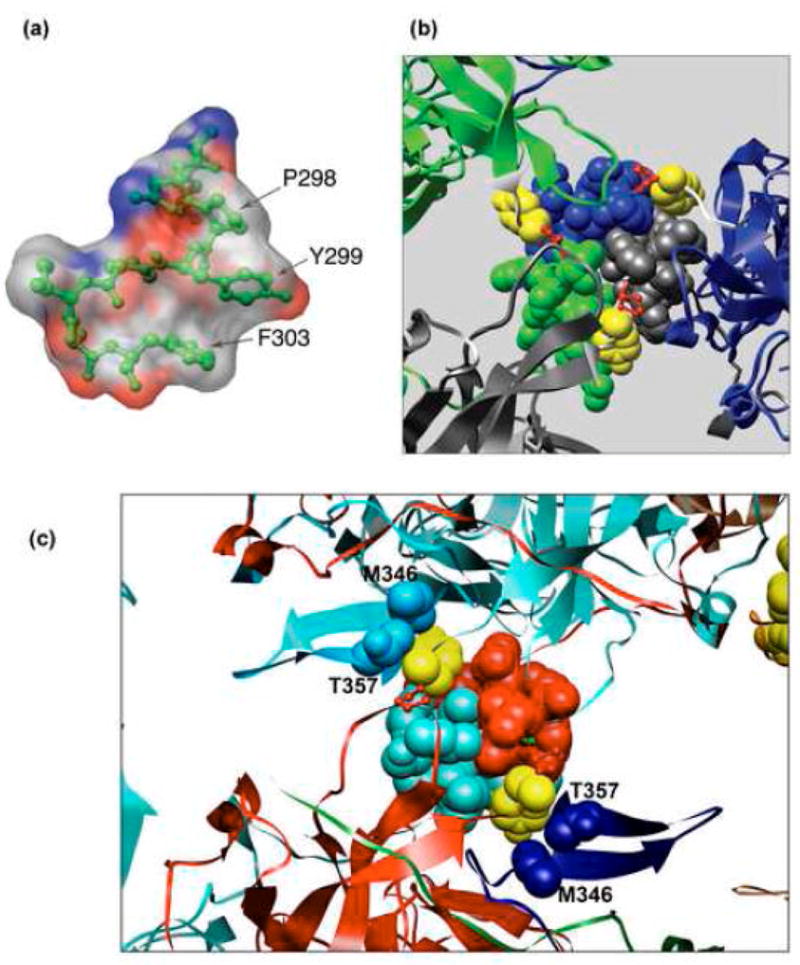
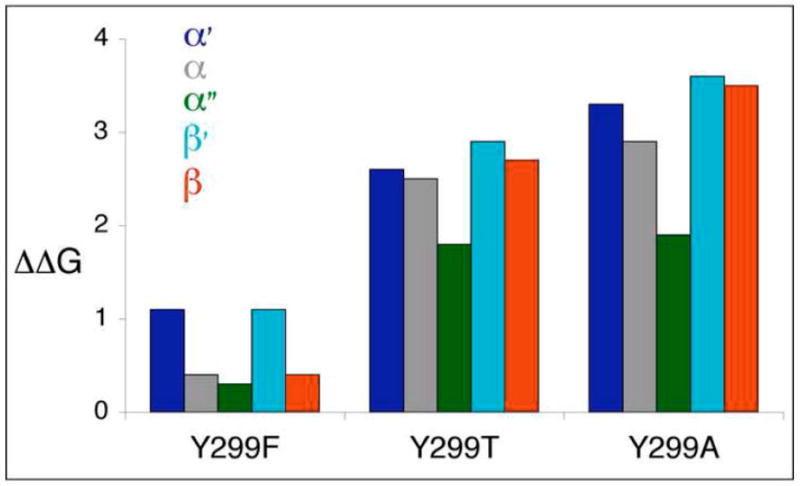
The simian virus 40 (SV40) outer shell is composed of 72 pentamers of VP1. The core of the VP1 monomer is a beta-barrel with jelly-roll topology and extending N- and C-terminal arms. A pentapeptide hinge, KNPYP, tethers the C-arm to the VP1 beta-barrel core. The five C-arms that extend from each pentamer insert into the neighbouring pentamers, tying them together through different types of interactions. In the mature virion, this element adopts either of six conformations according to their location in the capsid. We found that the hinge is conserved among 16 members of the Polyomaviridae, attesting to its importance in capsid assembly and/or structure. We have used site-directed mutagenesis to gain an understanding into the structural requirements of this element: Y299 was changed to A, F, and T, and P300 to A and G. The mutants showed reduction in viability to varying degrees. Unexpectedly, assembly was reduced only to a small extent. However, the data showed that the mutants were highly unstable. The largest effect was observed for mutations of P300, indicating a role of the proline in the virion structure. P300G was more unstable than P300A, indicating a requirement for rigidity of the pentapeptide hinge. Y299T and Y299A were more defective in viability than Y299F, highlighting the importance of an aromatic ring at this position. Structural inspection showed that this aromatic ring contacts C-arms of neighbouring pentamers. Computational modelling predicted loss of stability of the Y mutants in concordance with the experimental results. This study provides insights into the structural details of the pentapeptide hinge that are responsible for capsid stability.
Figures
Similar articles
-
Simian virus 40 Vp1 DNA-binding domain is functionally separable from the overlapping nuclear localization signal and is required for effective virion formation and full viability.J Virol. 2001 Aug;75(16):7321-9. doi: 10.1128/JVI.75.16.7321-7329.2001. J Virol. 2001. PMID: 11462004 Free PMC article.
-
Structure of simian virus 40 at 3.8-A resolution.Nature. 1991 Nov 28;354(6351):278-84. doi: 10.1038/354278a0. Nature. 1991. PMID: 1659663
-
Role of simian virus 40 Vp1 cysteines in virion infectivity.J Virol. 2000 Dec;74(23):11388-93. doi: 10.1128/jvi.74.23.11388-11393.2000. J Virol. 2000. PMID: 11070039 Free PMC article.
-
Simian virus 40 chromatin interaction with the capsid proteins.J Biomol Struct Dyn. 1983 Dec;1(3):689-704. doi: 10.1080/07391102.1983.10507475. J Biomol Struct Dyn. 1983. PMID: 6101085 Review.
-
Host range analysis of simian virus 40, BK virus and chimaeric SV40/BKV: relative expression of large T-antigen and Vp1 in infected and transformed cells.Dev Biol Stand. 1998;94:191-205. Dev Biol Stand. 1998. PMID: 9776240 Review.
Cited by
-
Discovery of novel druggable pockets on polyomavirus VP1 through crystallographic fragment-based screening to develop capsid assembly inhibitors.RSC Chem Biol. 2022 Apr 29;3(8):1013-1027. doi: 10.1039/d2cb00052k. eCollection 2022 Aug 3. RSC Chem Biol. 2022. PMID: 35974998 Free PMC article.
-
Assembly and Stability of Simian Virus 40 Polymorphs.ACS Nano. 2020 Apr 28;14(4):4430-4443. doi: 10.1021/acsnano.9b10004. Epub 2020 Apr 2. ACS Nano. 2020. PMID: 32208635 Free PMC article.
-
Nucleocapsid Assembly of Baculoviruses.Viruses. 2019 Jul 1;11(7):595. doi: 10.3390/v11070595. Viruses. 2019. PMID: 31266177 Free PMC article. Review.
-
Evidence of the Mechanism by Which Polyomaviruses Exploit the Extracellular Vesicle Delivery System during Infection.Viruses. 2020 May 27;12(6):585. doi: 10.3390/v12060585. Viruses. 2020. PMID: 32471033 Free PMC article. Review.
-
Scaffold properties are a key determinant of the size and shape of self-assembled virus-derived particles.ACS Chem Biol. 2013 Dec 20;8(12):2753-61. doi: 10.1021/cb4005518. Epub 2013 Oct 23. ACS Chem Biol. 2013. PMID: 24093474 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
