

Regulation of C-type lectin antimicrobial activity by a flexible N-terminal prosegment
- PMID: 19095652
- PMCID: PMC2643518
- DOI: 10.1074/jbc.M808077200
Regulation of C-type lectin antimicrobial activity by a flexible N-terminal prosegment
Abstract
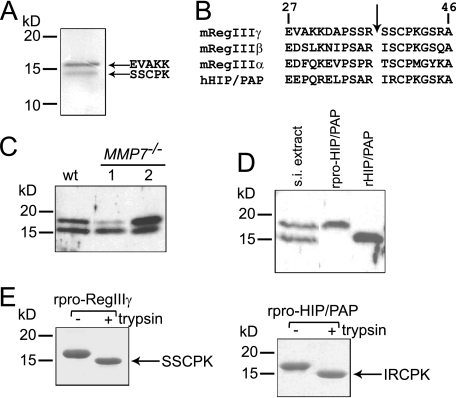
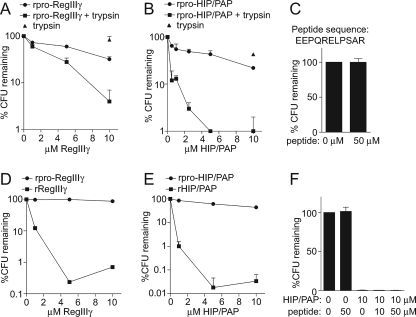
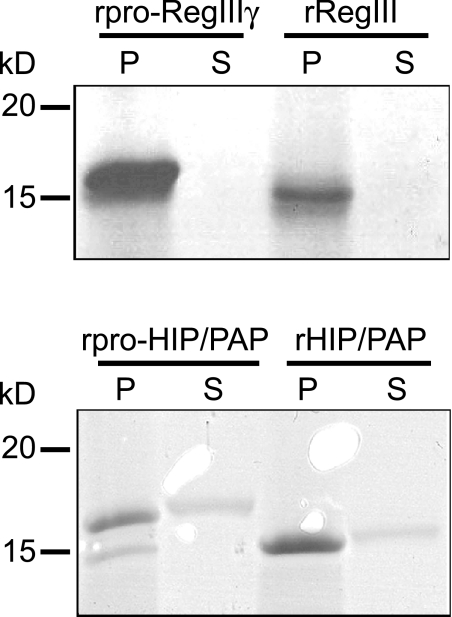
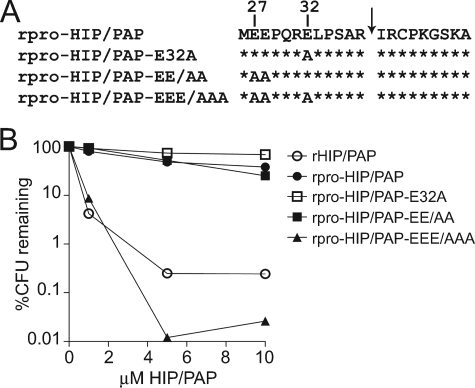
Members of the RegIII family of intestinal C-type lectins are directly antibacterial proteins that play a vital role in maintaining host-bacterial homeostasis in the mammalian gut, yet little is known about the mechanisms that regulate their biological activity. Here we show that the antibacterial activities of mouse RegIIIgamma and its human ortholog, HIP/PAP, are tightly controlled by an inhibitory N-terminal prosegment that is removed by trypsin in vivo. NMR spectroscopy revealed a high degree of conformational flexibility in the HIP/PAP inhibitory prosegment, and mutation of either acidic prosegment residues or basic core protein residues disrupted prosegment inhibitory activity. NMR analyses of pro-HIP/PAP variants revealed distinctive colinear backbone amide chemical shift changes that correlated with antibacterial activity, suggesting that prosegment-HIP/PAP interactions are linked to a two-state conformational switch between biologically active and inactive protein states. These findings reveal a novel regulatory mechanism governing C-type lectin biological function and yield new insight into the control of intestinal innate immunity.
Figures
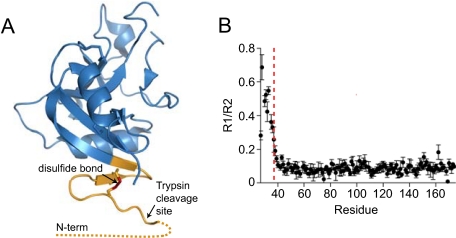
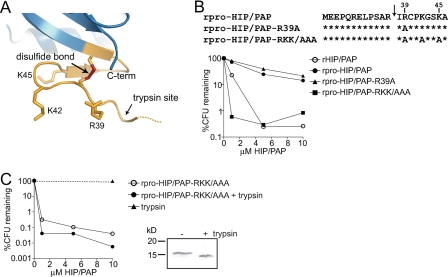
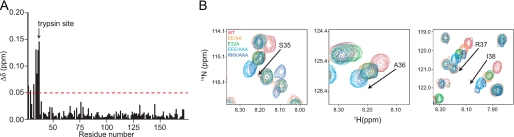
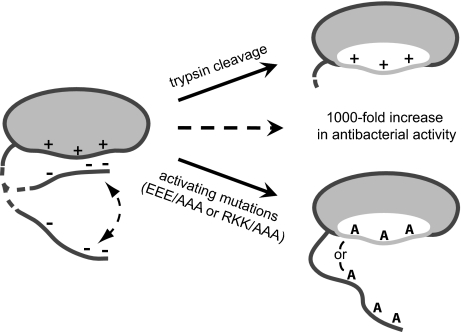
References
-
- Wilson, C. L., Ouellette, A. J., Satchell, D. P., Ayabe, T., Lopez-Boado, Y. S., Stratman, J. L., Hultgren, S. J., Matrisian, L. M., and Parks, W. C. (1999) Science 286 113–117 - PubMed
-
- Salzman, N. H., Ghosh, D., Huttner, K. M., Paterson, Y., and Bevins, C. L. (2003) Nature 422 522–526 - PubMed
-
- Salzman, N. H., Underwood, M. A., and Bevins, C. L. (2007) Semin. Immunol. 19 70–83 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
