

Dependence of bacterial chemotaxis on gradient shape and adaptation rate
- PMID: 19096502
- PMCID: PMC2588534
- DOI: 10.1371/journal.pcbi.1000242
Dependence of bacterial chemotaxis on gradient shape and adaptation rate
Abstract
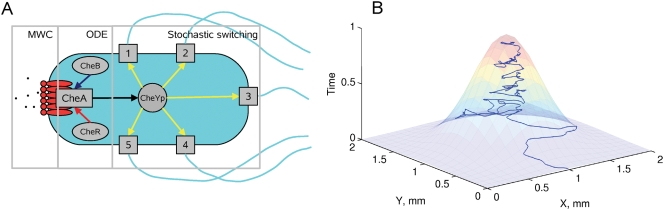
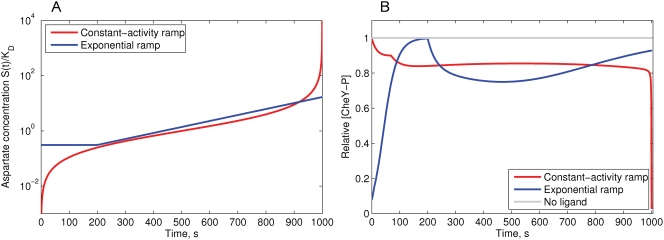
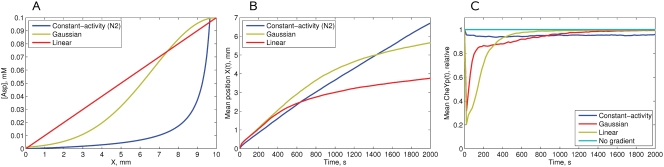
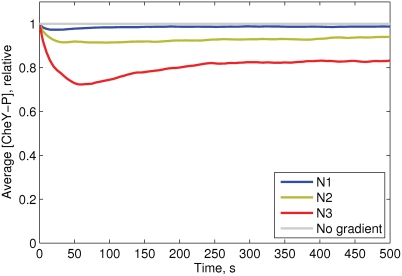
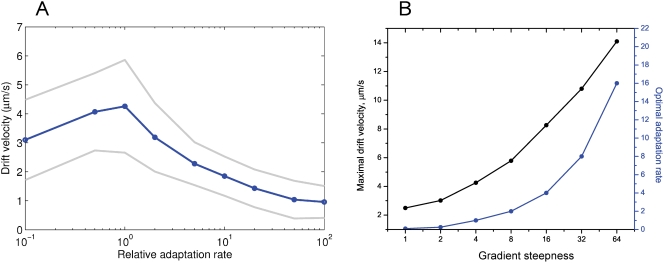
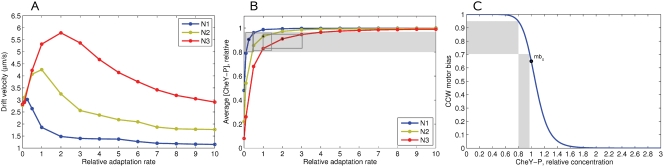
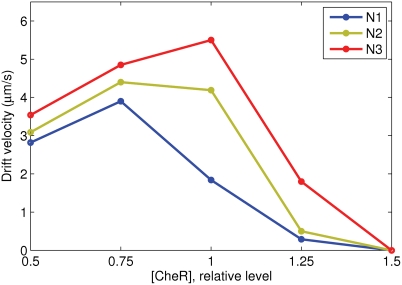
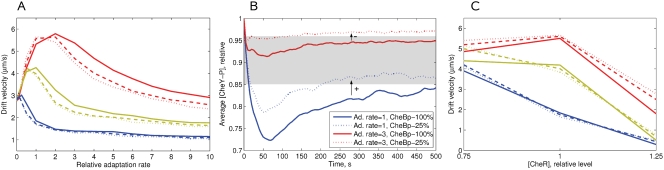
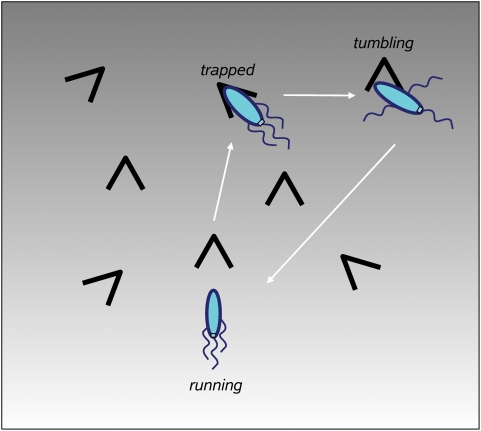
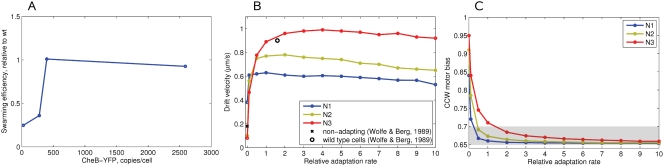
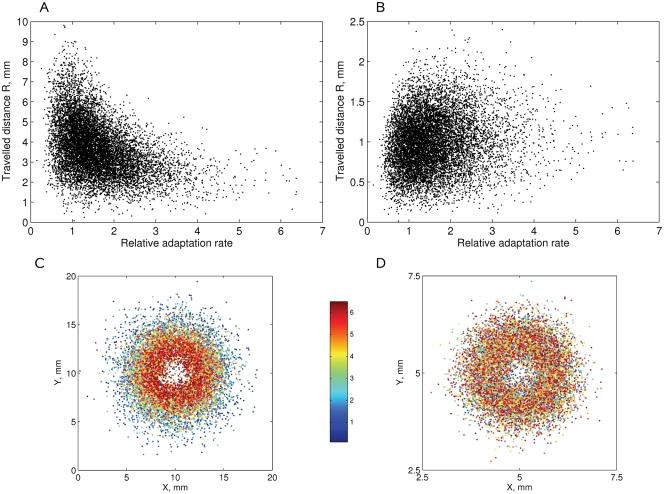
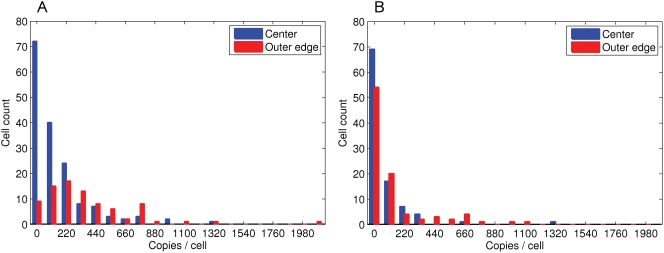
Simulation of cellular behavior on multiple scales requires models that are sufficiently detailed to capture central intracellular processes but at the same time enable the simulation of entire cell populations in a computationally cheap way. In this paper we present RapidCell, a hybrid model of chemotactic Escherichia coli that combines the Monod-Wyman-Changeux signal processing by mixed chemoreceptor clusters, the adaptation dynamics described by ordinary differential equations, and a detailed model of cell tumbling. Our model dramatically reduces computational costs and allows the highly efficient simulation of E. coli chemotaxis. We use the model to investigate chemotaxis in different gradients, and suggest a new, constant-activity type of gradient to systematically study chemotactic behavior of virtual bacteria. Using the unique properties of this gradient, we show that optimal chemotaxis is observed in a narrow range of CheA kinase activity, where concentration of the response regulator CheY-P falls into the operating range of flagellar motors. Our simulations also confirm that the CheB phosphorylation feedback improves chemotactic efficiency by shifting the average CheY-P concentration to fit the motor operating range. Our results suggest that in liquid media the variability in adaptation times among cells may be evolutionary favorable to ensure coexistence of subpopulations that will be optimally tactic in different gradients. However, in a porous medium (agar) such variability appears to be less important, because agar structure poses mainly negative selection against subpopulations with low levels of adaptation enzymes. RapidCell is available from the authors upon request.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Quantitative modeling of Escherichia coli chemotactic motion in environments varying in space and time.PLoS Comput Biol. 2010 Apr 8;6(4):e1000735. doi: 10.1371/journal.pcbi.1000735. PLoS Comput Biol. 2010. PMID: 20386737 Free PMC article.
-
The Role of Adaptation in Bacterial Speed Races.PLoS Comput Biol. 2016 Jun 3;12(6):e1004974. doi: 10.1371/journal.pcbi.1004974. eCollection 2016 Jun. PLoS Comput Biol. 2016. PMID: 27257812 Free PMC article.
-
Robustness analysis of the E. coli chemosensory system to perturbations in chemoattractant concentrations.Bioinformatics. 2007 Apr 1;23(7):875-81. doi: 10.1093/bioinformatics/btm028. Epub 2007 Jan 31. Bioinformatics. 2007. PMID: 17267424
-
Bacillus subtilis chemotaxis: a deviation from the Escherichia coli paradigm.Mol Microbiol. 1992 Jan;6(1):23-8. doi: 10.1111/j.1365-2958.1992.tb00833.x. Mol Microbiol. 1992. PMID: 1738311 Review.
-
Receptor clustering and signal processing in E. coli chemotaxis.Trends Microbiol. 2004 Dec;12(12):569-76. doi: 10.1016/j.tim.2004.10.003. Trends Microbiol. 2004. PMID: 15539117 Review.
Cited by
-
Pathway-based mean-field model for Escherichia coli chemotaxis.Phys Rev Lett. 2012 Jul 27;109(4):048101. doi: 10.1103/PhysRevLett.109.048101. Epub 2012 Jul 23. Phys Rev Lett. 2012. PMID: 23006109 Free PMC article.
-
Chemotactic response and adaptation dynamics in Escherichia coli.PLoS Comput Biol. 2010 May 20;6(5):e1000784. doi: 10.1371/journal.pcbi.1000784. PLoS Comput Biol. 2010. PMID: 20502674 Free PMC article.
-
Optimal chemotactic responses in stochastic environments.PLoS One. 2017 Jun 23;12(6):e0179111. doi: 10.1371/journal.pone.0179111. eCollection 2017. PLoS One. 2017. PMID: 28644830 Free PMC article.
-
Drift and Behavior of E. coli Cells.Biophys J. 2017 Dec 5;113(11):2321-2325. doi: 10.1016/j.bpj.2017.09.031. Epub 2017 Oct 27. Biophys J. 2017. PMID: 29111155 Free PMC article.
-
A hydrodynamic-stochastic model of chemotactic ciliated microorganisms.Eur Phys J E Soft Matter. 2019 Feb 21;42(2):20. doi: 10.1140/epje/i2019-11780-4. Eur Phys J E Soft Matter. 2019. PMID: 30788619
References
-
- Kennedy MJ. Role of motility, chemotaxis, and adhesion in microbial ecology. Ann N Y Acad Sci. 1987;506:260–273. - PubMed
-
- Pilgram WK, Williams FD. Survival value of chemotaxis in mixed cultures. Can J Microbiol. 1976;22:1771–1773. - PubMed
-
- Freter R, O'Brien PC, Halstead SA. Adhesion and chemotaxis as determinants of bacterial association with mucosal surfaces. Adv Exp Med Biol. 1978;107:429–437. - PubMed
-
- Lauffenburger D. Quantitative studies of bacterial chemotaxis and microbial population dynamics. Microb Ecol. 1991;22:175–185. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
