

Knock-out models reveal new aquaporin functions
- PMID: 19096787
- PMCID: PMC3595129
- DOI: 10.1007/978-3-540-79885-9_18
Knock-out models reveal new aquaporin functions
Abstract
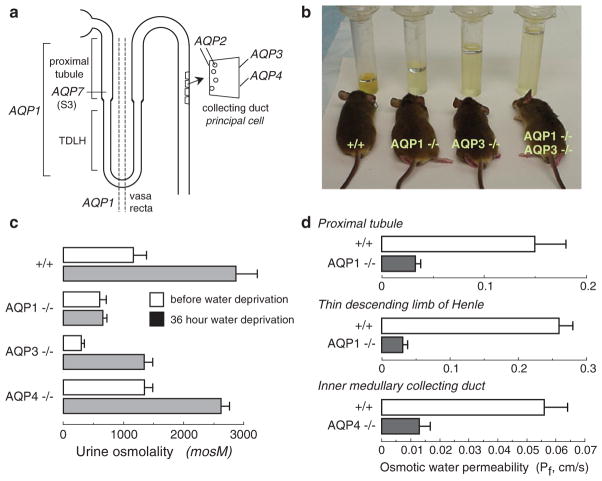
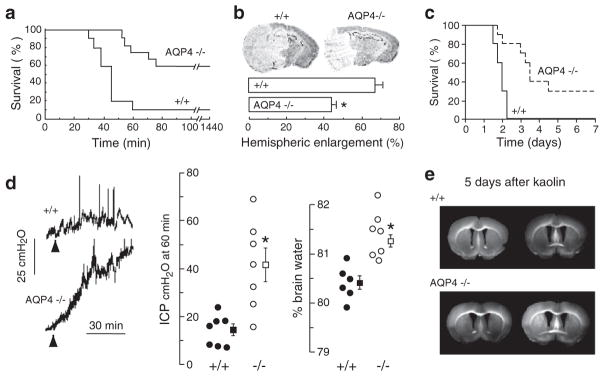
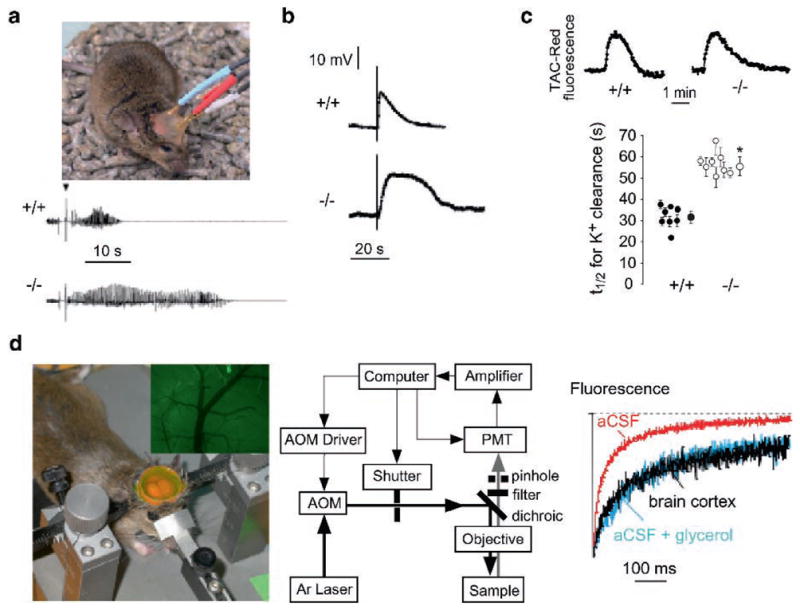
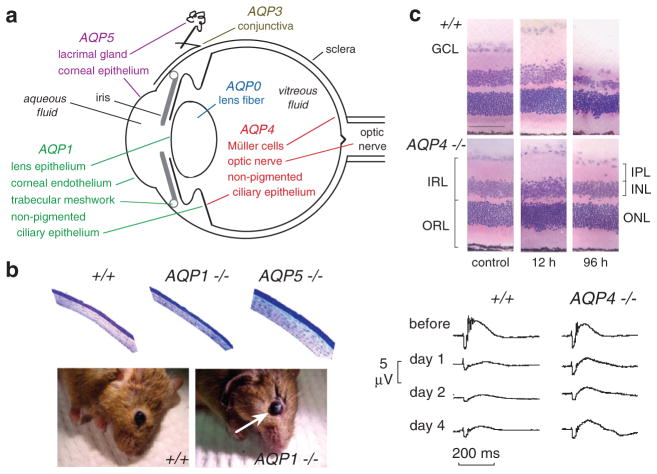
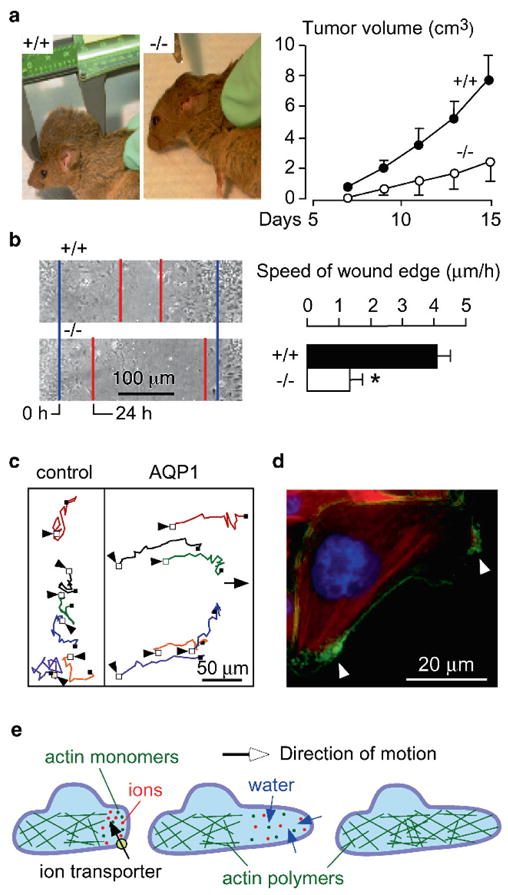
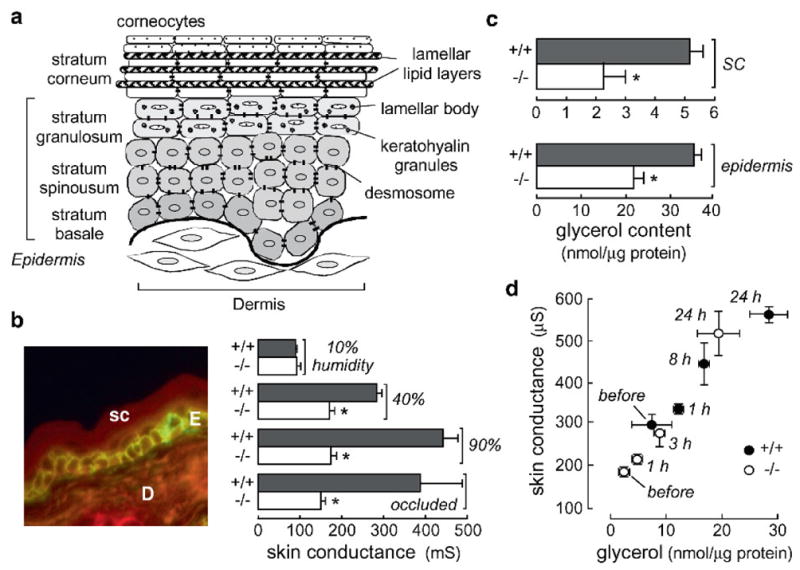
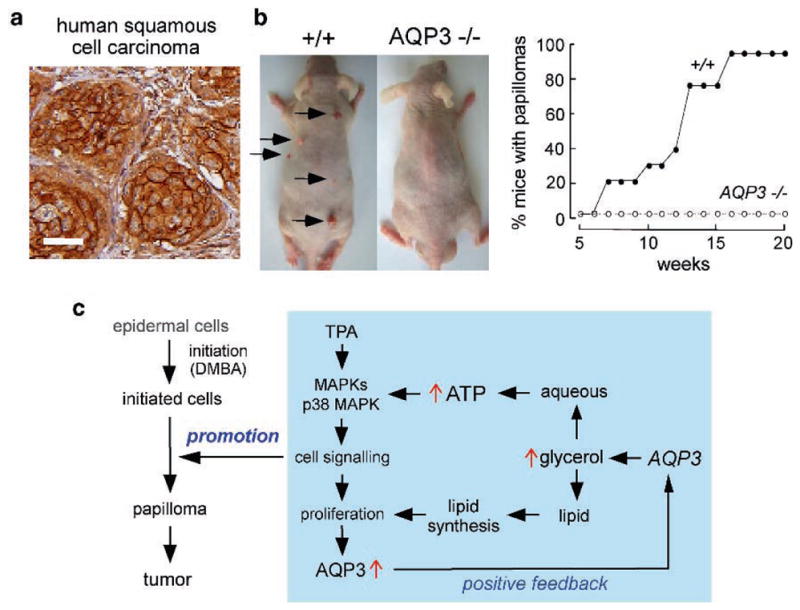
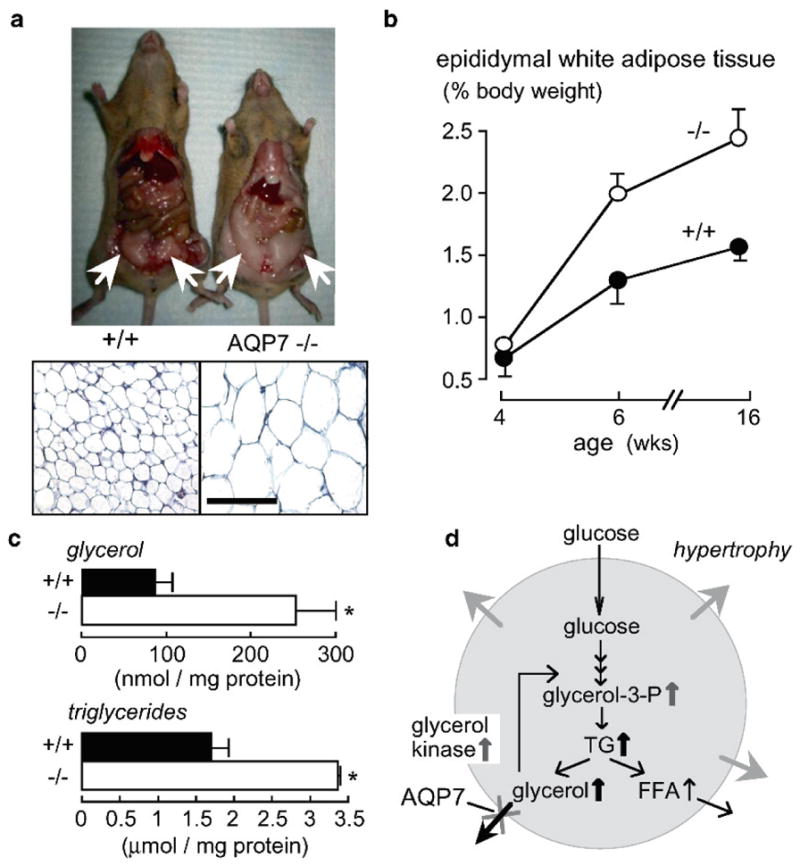
Knockout mice have been informative in the discovery of unexpected biological functions of aquaporins. Knockout mice have confirmed the predicted roles of aquaporins in transepithelial fluid transport, as in the urinary concentrating mechanism and glandular fluid secretion. A less obvious, though predictable role of aquaporins is in tissue swelling under stress, as in the brain in stroke, tumor and infection. Phenotype analysis of aquaporin knockout mice has revealed several unexpected cellular roles of aquaporins whose mechanisms are being elucidated. Aquaporins facilitate cell migration, as seen in aquaporin-dependent tumor angiogenesis and tumor metastasis, by a mechanism that may involve facilitated water transport in lamellipodia of migrating cells. The ' aquaglyceroporins', aquaporins that transport both glycerol and water, regulate glycerol content in epidermis, fat and other tissues, and lead to a multiplicity of interesting consequences of gene disruption including dry skin, resistance to skin carcinogenesis, impaired cell proliferation and altered fat metabolism. An even more surprising role of a mammalian aquaporin is in neural signal transduction in the central nervous system. The many roles of aquaporins might be exploited for clinical benefit by modulation of aquaporin expression/function - as diuretics, and in the treatment of brain swelling, glaucoma, epilepsy, obesity and cancer.
Figures
Similar articles
-
Mammalian aquaporins: diverse physiological roles and potential clinical significance.Expert Rev Mol Med. 2008 May 16;10:e13. doi: 10.1017/S1462399408000690. Expert Rev Mol Med. 2008. PMID: 18482462 Free PMC article. Review.
-
Aquaporins: translating bench research to human disease.J Exp Biol. 2009 Jun;212(Pt 11):1707-15. doi: 10.1242/jeb.024125. J Exp Biol. 2009. PMID: 19448080 Free PMC article. Review.
-
More than just water channels: unexpected cellular roles of aquaporins.J Cell Sci. 2005 Aug 1;118(Pt 15):3225-32. doi: 10.1242/jcs.02519. J Cell Sci. 2005. PMID: 16079275 Review.
-
Non-Transport Functions of Aquaporins.Adv Exp Med Biol. 2023;1398:65-80. doi: 10.1007/978-981-19-7415-1_5. Adv Exp Med Biol. 2023. PMID: 36717487
-
Aquaporins as potential drug targets.Acta Pharmacol Sin. 2006 Apr;27(4):395-401. doi: 10.1111/j.1745-7254.2006.00318.x. Acta Pharmacol Sin. 2006. PMID: 16539837 Review.
Cited by
-
Important functions and molecular mechanisms of aquaporins family on respiratory diseases: potential translational values.J Cancer. 2024 Oct 7;15(18):6073-6085. doi: 10.7150/jca.98829. eCollection 2024. J Cancer. 2024. PMID: 39440058 Free PMC article. Review.
-
Aquaporins in the adult mouse submandibular and sublingual salivary glands.J Mol Histol. 2014 Feb;45(1):69-80. doi: 10.1007/s10735-013-9526-3. Epub 2013 Jul 24. J Mol Histol. 2014. PMID: 23880985
-
Recovery of Water Homeostasis in Adenine-Induced Kidney Disease Is Mediated by Increased AQP2 Membrane Targeting.Int J Mol Sci. 2024 Mar 19;25(6):3447. doi: 10.3390/ijms25063447. Int J Mol Sci. 2024. PMID: 38542420 Free PMC article.
-
Aquaporin 1 knockdown inhibits triple-negative breast cancer cell proliferation and invasion in vitro and in vivo.Oncol Lett. 2021 Jun;21(6):437. doi: 10.3892/ol.2021.12698. Epub 2021 Apr 1. Oncol Lett. 2021. PMID: 33868475 Free PMC article.
-
Distribution of intraperitoneally administered deuterium-labeled water in aquaporin-4-knockout mouse brain after middle cerebral artery occlusion.Front Neurosci. 2023 Jan 6;16:1071272. doi: 10.3389/fnins.2022.1071272. eCollection 2022. Front Neurosci. 2023. PMID: 36685250 Free PMC article.
References
-
- Amiry-Moghaddam M, Otsuka T, Hurn PD, Traystman RJ, Haug FM, Froehner SC, Adams ME, Neely JD, Agre P, Ottersen OP, Bhardwaj A. An alpha-syntrophin-dependent pool of AQP4 in astroglial end-feet confers bidirectional water flow between blood and brain. Proc Natl Acad Sci U S A. 2003a;100:2106–2111. - PMC - PubMed
-
- Amiry-Moghaddam M, Williamson A, Palomba M, Eid T, de Lanerolle NC, Nagelhus EA, Adams ME, Froehner SC, Agre P, Ottersen OP. Delayed K+ clearance associated with aquaporin-4 mislocalization: phenotypic defects in brains of alpha-syntrophin-null mice. Proc Natl Acad Sci U S A. 2003b;100:13615–13620. - PMC - PubMed
-
- Auguste KI, Jin S, Uchida K, Yan D, Manley GT, Papadopoulos MC, Verkman AS. Greatly impaired migration of implanted aquaporin-4-deficient astroglial cells in mouse brain toward a site of injury. FASEB J. 2007;21:108–116. - PubMed
-
- Berry V, Francis P, Kaushal S, Moore A, Bhattacharya S. Missense mutations in MIP underlie autosomal dominant ‘polymorphic’ and lamellar cataracts linked to 12q. Nat Genet. 2000;25:15–17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HL73856/HL/NHLBI NIH HHS/United States
- R01 EY013574/EY/NEI NIH HHS/United States
- R01 EB000415/EB/NIBIB NIH HHS/United States
- R01 DK035124/DK/NIDDK NIH HHS/United States
- EY13574/EY/NEI NIH HHS/United States
- DK35124/DK/NIDDK NIH HHS/United States
- EB00415/EB/NIBIB NIH HHS/United States
- HL59198/HL/NHLBI NIH HHS/United States
- R01 HL073856/HL/NHLBI NIH HHS/United States
- P30 DK072517/DK/NIDDK NIH HHS/United States
- DK72517/DK/NIDDK NIH HHS/United States
- R01 HL059198/HL/NHLBI NIH HHS/United States
- R37 DK035124/DK/NIDDK NIH HHS/United States
- R37 EB000415/EB/NIBIB NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
