

The human complement fragment receptor, C5L2, is a recycling decoy receptor
- PMID: 19100624
- PMCID: PMC2697321
- DOI: 10.1016/j.molimm.2008.11.001
The human complement fragment receptor, C5L2, is a recycling decoy receptor
Abstract
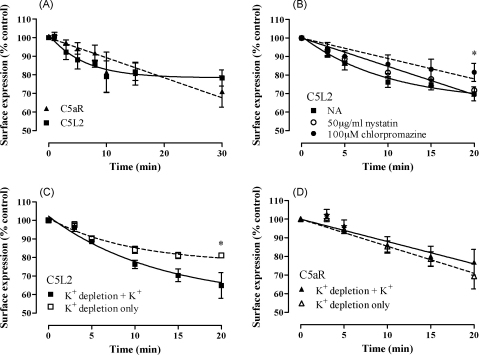
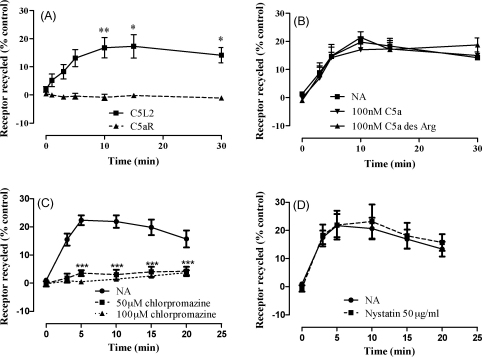
C5L2 is a 7 transmembrane domain receptor for complement fragment C5a that, unlike the classical C5a receptor, C5aR, does not couple to G proteins. However, in mice where C5L2 has been deleted, the response to C5a is altered, suggesting that C5L2 may have a signaling function. In order to investigate whether human C5L2 also has some capacity to transduce signals, we have attempted to produce a signaling competent form of human C5L2 by inserting C5aR sequences at three key G protein activation motifs. However, we detected neither an intracellular Ca(2+) response nor beta-arrestin redistribution in mutated C5L2, suggesting that the potential for G protein coupling is completely absent in this receptor and that, in humans, C5L2 may have functions that are unrelated to signaling. In confirmation of this, we detected constitutive ligand-independent internalization of C5L2 that resulted in the rapid accumulation of C5a and its stable metabolite, C5a des Arg, within the cell with only a small net change in cell surface receptor levels. Internalization was found to be through a clathrin-dependent mechanism that led to the retention and, in cells natively expressing C5L2, the degradation of the ligand within an intracellular compartment. In contrast, the classical C5a receptor, C5aR, internalized ligand much more slowly and a majority of this ligand was released back into the extracellular environment in an apparently undegraded form. These data suggest that a major function of human C5L2 is to remove active complement fragments from the extracellular environment.
Figures









References
-
- Barak L.S., Tiberi M., Freedman N.J., Kwatra M.M., Lefkowitz R.J., Caron M.G. A highly conserved tyrosine residue in G protein-coupled receptors is required for agonist-mediated beta 2-adrenergic receptor sequestration. J. Biol. Chem. 1994;269:2790–2795. - PubMed
-
- Bonecchi R., Borroni E.M., Anselmo A., Doni A., Savino B., Mirolo M., Fabbri M., Jala V.R., Haribabu B., Mantovani A., Locati M. Regulation of D6 chemokine scavenging activity by ligand and Rab11-dependent surface upregulation. Blood. 2008;112:493–503. - PubMed
-
- Borroni E.M., Buracchi C., de la Torre Y.M., Galliera E., Vecchi A., Bonecchi R., Mantovani A., Locati M. The chemoattractant decoy receptor D6 as a negative regulator of inflammatory responses. Biochem. Soc. Trans. 2006;34:1014–1017. - PubMed
-
- Boulay F., Mery L., Tardif M., Brouchon L., Vignais P. Expression cloning of a receptor for C5a anaphylatoxin on differentiated HL-60 cells. Biochemistry. 1991;30:2993–2999. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
