

Optimization of in vivo activity of a bifunctional homing endonuclease and maturase reverses evolutionary degradation
- PMID: 19103658
- PMCID: PMC2647301
- DOI: 10.1093/nar/gkn1007
Optimization of in vivo activity of a bifunctional homing endonuclease and maturase reverses evolutionary degradation
Abstract
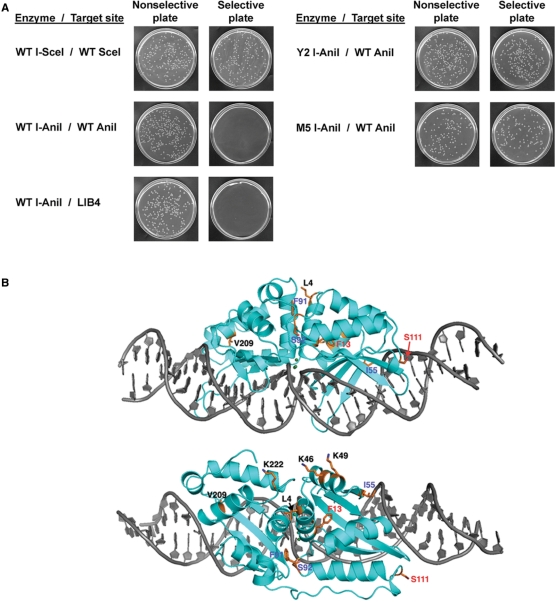
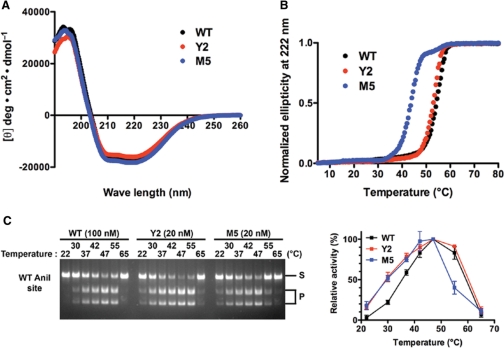
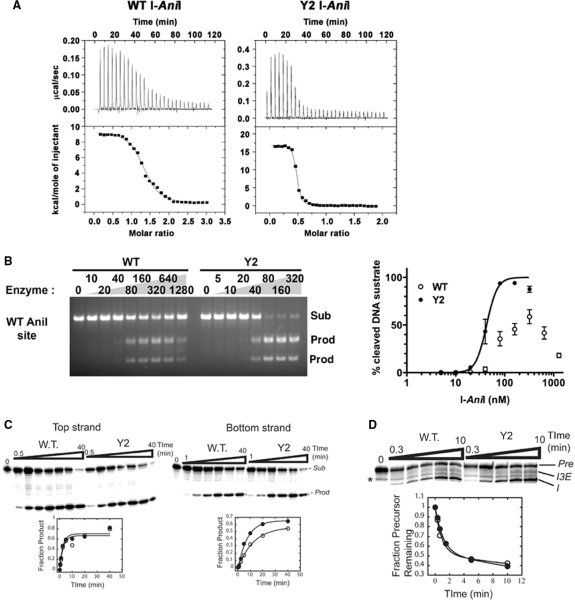
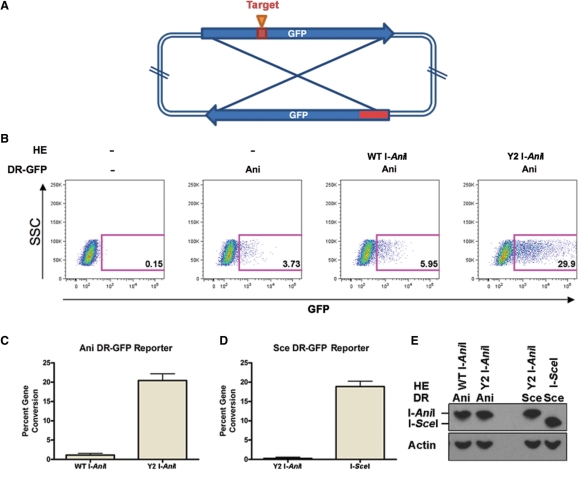
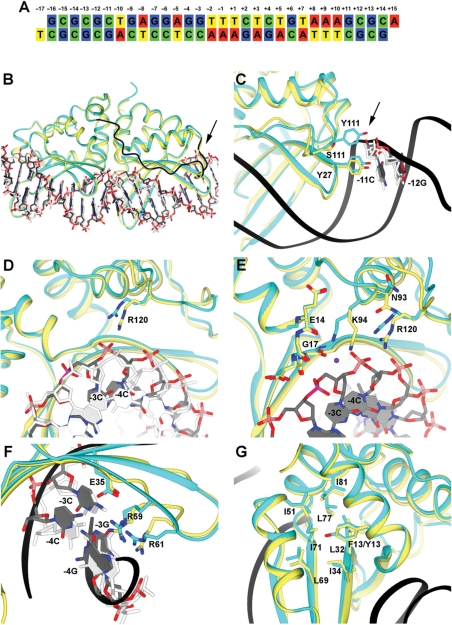
The LAGLIDADG homing endonuclease (LHE) I-AniI has adopted an extremely efficient secondary RNA splicing activity that is beneficial to its host, balanced against inefficient DNA cleavage. A selection experiment identified point mutations in the enzyme that act synergistically to improve endonuclease activity. The amino-acid substitutions increase target affinity, alter the thermal cleavage profile and significantly increase targeted recombination in transfected cells. The RNA splicing activity is not affected by these mutations. The improvement in DNA cleavage activity is largely focused on one of the enzyme's two active sites, corresponding to a rearrangement of a lysine residue hypothesized to act as a general base. Most of the constructs isolated in the screen contain one or more mutations that revert an amino-acid identity to a residue found in one or more close homologues of I-AniI. This implies that mutations that have previously reduced the endonuclease activity of I-AniI are identified and reversed, sometimes in combination with additional 'artificial' mutations, to optimize its in vivo activity.
Figures

Similar articles
-
Functionally distinct nucleic acid binding sites for a group I intron encoded RNA maturase/DNA homing endonuclease.J Mol Biol. 2003 May 30;329(2):239-51. doi: 10.1016/s0022-2836(03)00426-1. J Mol Biol. 2003. PMID: 12758073
-
In vitro analysis of the relationship between endonuclease and maturase activities in the bi-functional group I intron-encoded protein, I-AniI.Eur J Biochem. 2003 Apr;270(7):1543-54. doi: 10.1046/j.1432-1033.2003.03518.x. Eur J Biochem. 2003. PMID: 12654010
-
Generation of a nicking enzyme that stimulates site-specific gene conversion from the I-AniI LAGLIDADG homing endonuclease.Proc Natl Acad Sci U S A. 2009 Mar 31;106(13):5099-104. doi: 10.1073/pnas.0810588106. Epub 2009 Mar 10. Proc Natl Acad Sci U S A. 2009. PMID: 19276110 Free PMC article.
-
Homing endonuclease structure and function.Q Rev Biophys. 2005 Feb;38(1):49-95. doi: 10.1017/S0033583505004063. Epub 2005 Dec 9. Q Rev Biophys. 2005. PMID: 16336743 Review.
-
Structural and functional characteristics of homing endonucleases.Crit Rev Biochem Mol Biol. 2003;38(3):199-248. doi: 10.1080/713609235. Crit Rev Biochem Mol Biol. 2003. PMID: 12870715 Review.
Cited by
-
Expanding LAGLIDADG endonuclease scaffold diversity by rapidly surveying evolutionary sequence space.Nucleic Acids Res. 2012 Jun;40(11):4954-64. doi: 10.1093/nar/gkr1303. Epub 2012 Feb 14. Nucleic Acids Res. 2012. PMID: 22334611 Free PMC article.
-
Homing endonucleases catalyze double-stranded DNA breaks and somatic transgene excision in Aedes aegypti.Insect Mol Biol. 2009 Oct;18(5):623-33. doi: 10.1111/j.1365-2583.2009.00905.x. Insect Mol Biol. 2009. PMID: 19754740 Free PMC article.
-
Progressive engineering of a homing endonuclease genome editing reagent for the murine X-linked immunodeficiency locus.Nucleic Acids Res. 2014 Jun;42(10):6463-75. doi: 10.1093/nar/gku224. Epub 2014 Mar 25. Nucleic Acids Res. 2014. PMID: 24682825 Free PMC article.
-
Exploitation of binding energy for catalysis and design.Nature. 2009 Oct 29;461(7268):1300-4. doi: 10.1038/nature08508. Nature. 2009. PMID: 19865174 Free PMC article.
-
Germline excision of transgenes in Aedes aegypti by homing endonucleases.Sci Rep. 2013;3:1603. doi: 10.1038/srep01603. Sci Rep. 2013. PMID: 23549343 Free PMC article.
References
-
- Stoddard BL. Homing endonuclease structure and function. Quarterly Reviews of Biophysics. 2005;38:49–95. - PubMed
-
- Jacquier A, Dujon B. An intron-encoded protein is active in a gene conversion process that spreads an intron into a mitochondrial gene. Cell. 1985;41:383–394. - PubMed
-
- Kostriken R, Strathern JN, Klar AJ, Hicks JB, Heffron F. A site-specific endonuclease essential for mating-type switching in Saccharomyces cerevisiae. Cell. 1983;35:167–174. - PubMed
-
- Colleaux L, d'Auriol L, Betermier M, Cottarel G, Jacquier A, Galibert F, Dujon B. Universal code equivalent of a yeast mitochondrial intron reading frame is expressed into E. coli as a specific double strand endonuclease. Cell. 1986;44:521–533. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
