

Neuronal oscillations and the rate-to-phase transform: mechanism, model and mutual information
- PMID: 19103680
- PMCID: PMC2669970
- DOI: 10.1113/jphysiol.2008.164111
Neuronal oscillations and the rate-to-phase transform: mechanism, model and mutual information
Abstract
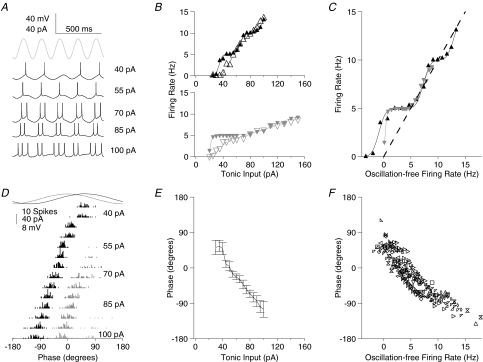
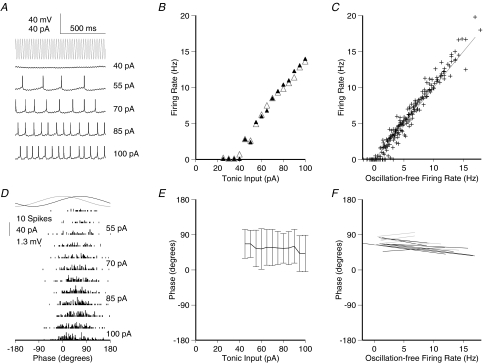
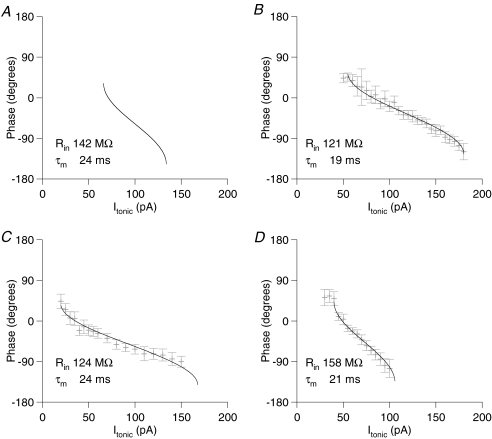
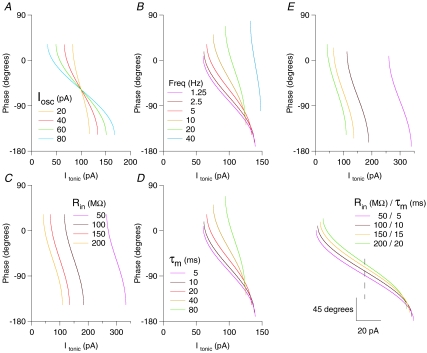
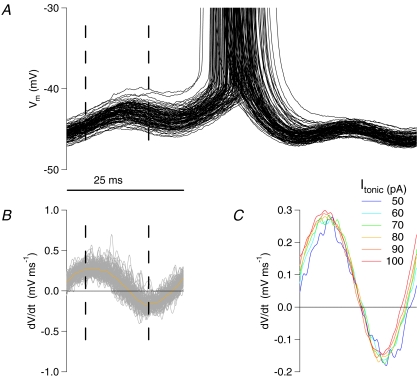
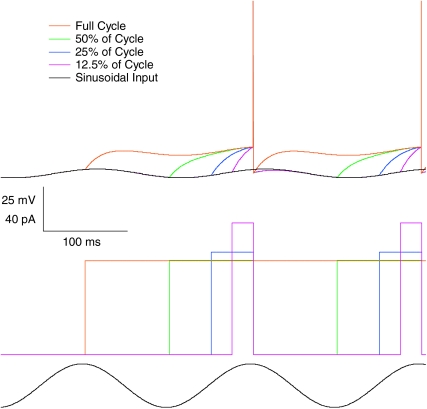
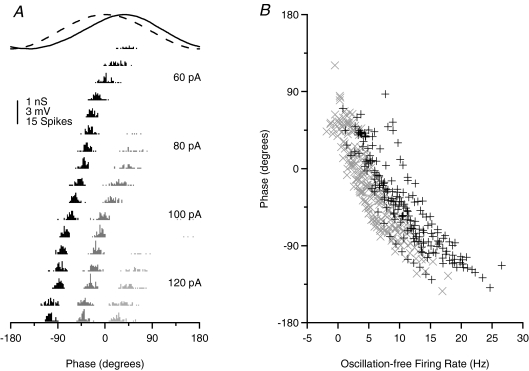
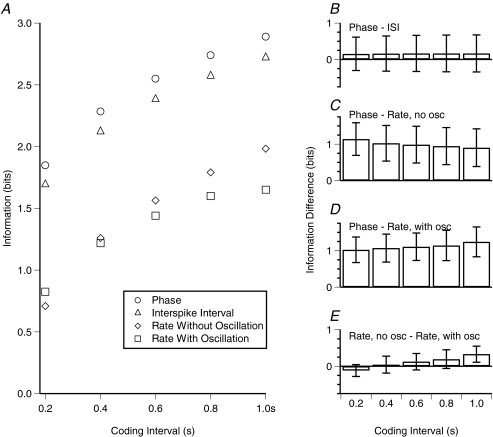
Theoretical and experimental studies suggest that oscillatory modes of processing play an important role in neuronal computations. One well supported idea is that the net excitatory input during oscillations will be reported in the phase of firing, a 'rate-to-phase transform', and that this transform might enable a temporal code. Here, we investigate the efficiency of this code at the level of fundamental single cell computations. We first develop a general framework for the understanding of the rate-to-phase transform as implemented by single neurons. Using whole cell patch-clamp recordings of rat hippocampal pyramidal neurons in vitro, we investigated the relationship between tonic excitation and phase of firing during simulated theta frequency (5 Hz) and gamma frequency (40 Hz) oscillations, over a range of physiological firing rates. During theta frequency oscillations, the phase of the first spike per cycle was a near-linear function of tonic excitation, advancing through a full 180 deg, from the peak to the trough of the oscillation cycle as excitation increased. In contrast, this relationship was not apparent for gamma oscillations, during which the phase of firing was virtually independent of the level of tonic excitatory input within the range of physiological firing rates. We show that a simple analytical model can substantially capture this behaviour, enabling generalization to other oscillatory states and cell types. The capacity of such a transform to encode information is limited by the temporal precision of neuronal activity. Using the data from our whole cell recordings, we calculated the information about the input available in the rate or phase of firing, and found the phase code to be significantly more efficient. Thus, temporal modes of processing can enable neuronal coding to be inherently more efficient, thereby allowing a reduction in processing time or in the number of neurons required.
Figures
Similar articles
-
Repetitive firing and oscillatory activity of pyramidal-like bursting neurons in the rat subiculum.Exp Brain Res. 1997 May;114(3):507-17. doi: 10.1007/pl00005660. Exp Brain Res. 1997. PMID: 9187287
-
Kv7/KCNQ channels control action potential phasing of pyramidal neurons during hippocampal gamma oscillations in vitro.J Neurosci. 2009 Oct 21;29(42):13353-64. doi: 10.1523/JNEUROSCI.1463-09.2009. J Neurosci. 2009. PMID: 19846723 Free PMC article.
-
Spike timing of distinct types of GABAergic interneuron during hippocampal gamma oscillations in vitro.J Neurosci. 2004 Oct 13;24(41):9127-37. doi: 10.1523/JNEUROSCI.2113-04.2004. J Neurosci. 2004. PMID: 15483131 Free PMC article.
-
The theta/gamma discrete phase code occuring during the hippocampal phase precession may be a more general brain coding scheme.Hippocampus. 2005;15(7):913-22. doi: 10.1002/hipo.20121. Hippocampus. 2005. PMID: 16161035 Review.
-
Cellular mechanisms of neuronal population oscillations in the hippocampus in vitro.Annu Rev Neurosci. 2004;27:247-78. doi: 10.1146/annurev.neuro.27.070203.144303. Annu Rev Neurosci. 2004. PMID: 15217333 Review.
Cited by
-
Theta oscillations optimize a speed-precision trade-off in phase coding neurons.PLoS Comput Biol. 2024 Dec 2;20(12):e1012628. doi: 10.1371/journal.pcbi.1012628. eCollection 2024 Dec. PLoS Comput Biol. 2024. PMID: 39621800 Free PMC article.
-
Physiological features of parvalbumin-expressing GABAergic interneurons contributing to high-frequency oscillations in the cerebral cortex.Curr Res Neurobiol. 2023 Dec 16;6:100121. doi: 10.1016/j.crneur.2023.100121. eCollection 2024. Curr Res Neurobiol. 2023. PMID: 38616956 Free PMC article. Review.
-
Phase Difference between Model Cortical Areas Determines Level of Information Transfer.Front Comput Neurosci. 2017 Feb 9;11:6. doi: 10.3389/fncom.2017.00006. eCollection 2017. Front Comput Neurosci. 2017. PMID: 28232796 Free PMC article.
-
Phase of firing as a local window for efficient neuronal computation: tonic and phasic mechanisms in the control of theta spike phase.Front Hum Neurosci. 2011 Feb 2;5:3. doi: 10.3389/fnhum.2011.00003. eCollection 2011. Front Hum Neurosci. 2011. PMID: 21344003 Free PMC article. No abstract available.
-
A model of intracellular θ phase precession dependent on intrinsic subthreshold membrane currents.J Neurosci. 2011 Aug 24;31(34):12282-96. doi: 10.1523/JNEUROSCI.0586-11.2011. J Neurosci. 2011. PMID: 21865471 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources