

Mechanism of a prototypical synthetic membrane-active antimicrobial: Efficient hole-punching via interaction with negative intrinsic curvature lipids
- PMID: 19106303
- PMCID: PMC2634941
- DOI: 10.1073/pnas.0806456105
Mechanism of a prototypical synthetic membrane-active antimicrobial: Efficient hole-punching via interaction with negative intrinsic curvature lipids
Abstract
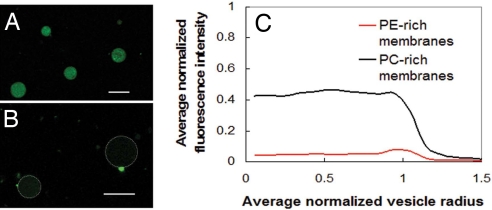
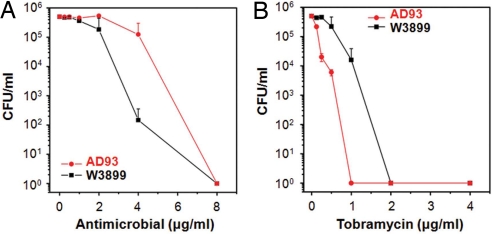
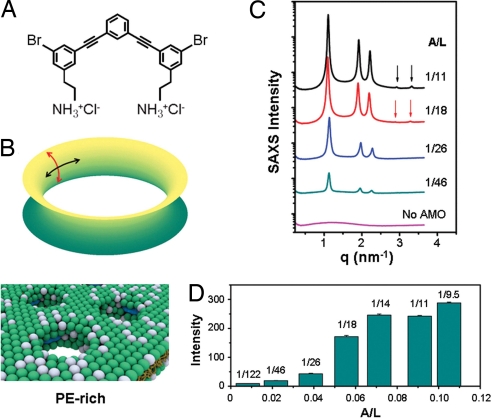
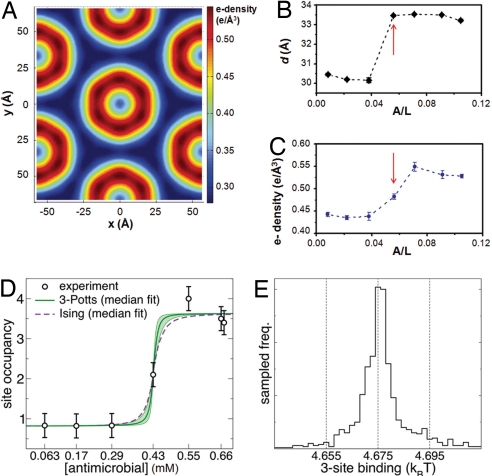
Phenylene ethynylenes comprise a prototypical class of synthetic antimicrobial compounds that mimic antimicrobial peptides produced by eukaryotes and have broad-spectrum antimicrobial activity. We show unambiguously that bacterial membrane permeation by these antimicrobials depends on the presence of negative intrinsic curvature lipids, such as phosphatidylethanolamine (PE) lipids, found in high concentrations within bacterial membranes. Plate-killing assays indicate that a PE-knockout mutant strain of Escherichia coli drastically out-survives the wild type against the membrane-active phenylene ethynylene antimicrobials, whereas the opposite is true when challenged with traditional metabolic antibiotics. That the PE deletion is a lethal mutation in normative environments suggests that resistant bacterial strains do not evolve because a lethal mutation is required to gain immunity. PE lipids allow efficient generation of negative curvature required for the circumferential barrel of an induced membrane pore; an inverted hexagonal H(II) phase, which consists of arrays of water channels, is induced by a small number of antimicrobial molecules. The estimated antimicrobial occupation in these water channels is nonlinear and jumps from approximately 1 to 3 per 4 nm of induced water channel length as the global antimicrobial concentration is increased. By comparing to exactly solvable 1D spin models for magnetic systems, we quantify the cooperativity of these antimicrobials.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Tomasz A. Multiple-antibiotic-resistant pathogenic bacteria – A report on the Rockefeller University workshop. N Engl J Med. 1994;330:1247–1251. - PubMed
-
- Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415:389–395. - PubMed
-
- Brogden KA. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat Rev Microbiol. 2005;3:238–250. - PubMed
-
- Hancock REW, Lehrer R. Cationic peptides: A new source of antibiotics. Trends Biotechnol. 1998;16:82–88. - PubMed
-
- Hancock REW, Sahl H-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat Biotech. 2006;24:1551–1557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
