

Stimulation of Toll-like receptor 2 in human platelets induces a thromboinflammatory response through activation of phosphoinositide 3-kinase
- PMID: 19106411
- PMCID: PMC2732983
- DOI: 10.1161/CIRCRESAHA.108.185785
Stimulation of Toll-like receptor 2 in human platelets induces a thromboinflammatory response through activation of phosphoinositide 3-kinase
Abstract
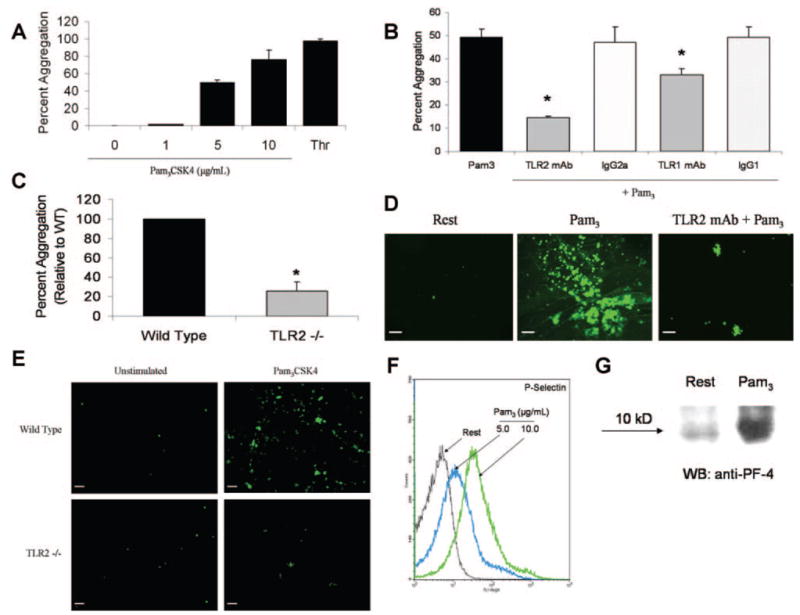
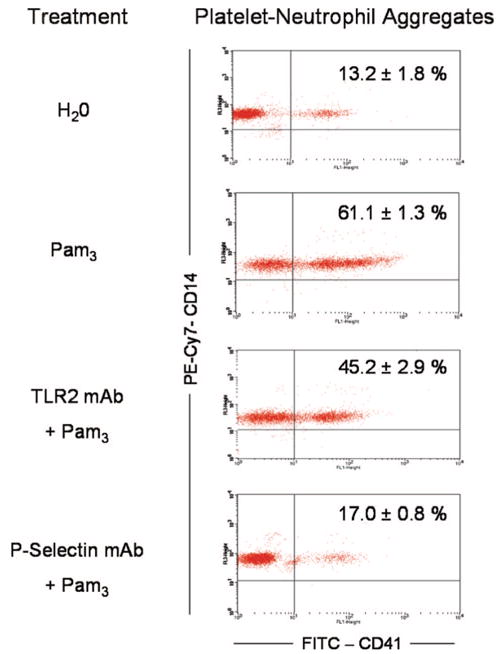
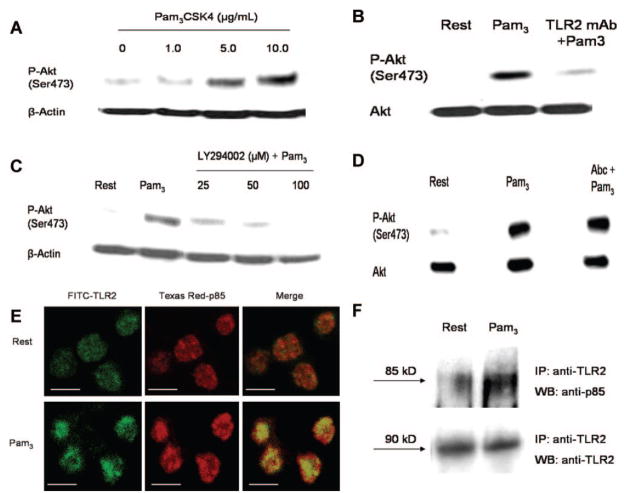
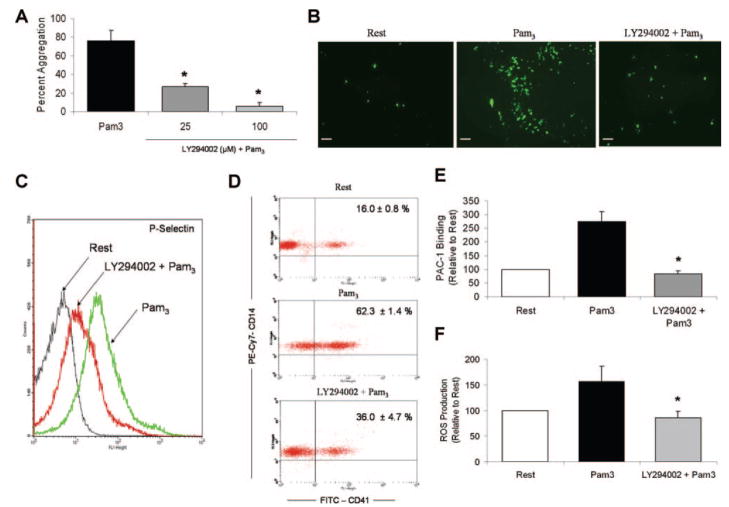
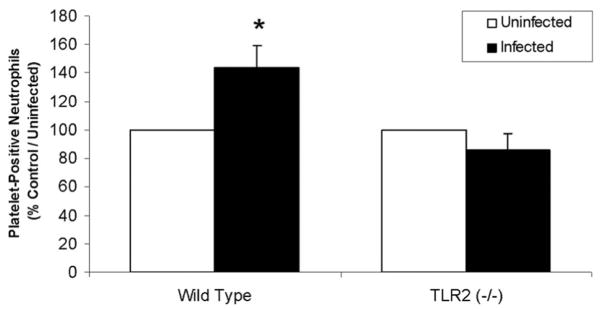
Cells of the innate immune system use Toll-like receptors (TLRs) to initiate the proinflammatory response to microbial infection. Recent studies have shown acute infections are associated with a transient increase in the risk of vascular thrombotic events. Although platelets play a central role in acute thrombosis and accumulating evidence demonstrates their role in inflammation and innate immunity, investigations into the expression and functionality of platelet TLRs have been limited. In the present study, we demonstrate that human platelets express TLR2, TLR1, and TLR6. Incubation of isolated platelets with Pam(3)CSK4, a synthetic TLR2/TLR1 agonist, directly induced platelet aggregation and adhesion to collagen. These functional responses were inhibited in TLR2-deficient mice and, in human platelets, by pretreatment with TLR2-blocking antibody. Stimulation of platelet TLR2 also increased P-selectin surface expression, activation of integrin alpha(IIb)beta(3), generation of reactive oxygen species, and, in human whole blood, formation of platelet-neutrophil heterotypic aggregates. TLR2 stimulation also activated the phosphoinositide 3-kinase (PI3-K)/Akt signaling pathway in platelets, and inhibition of PI3-K significantly reduced Pam(3)CSK4-induced platelet responses. In vivo challenge with live Porphyromonas gingivalis, a Gram-negative pathogenic bacterium that uses TLR2 for innate immune signaling, also induced significant formation of platelet-neutrophil aggregates in wild-type but not TLR2-deficient mice. Together, these data provide the first demonstration that human platelets express functional TLR2 capable of recognizing bacterial components and activating the platelet thrombotic and/or inflammatory pathways. This work substantiates the role of platelets in the immune and inflammatory response and suggests a mechanism by which bacteria could directly activate platelets.
Figures
References
-
- Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4:499–511. - PubMed
-
- Iwasaki A, Medzhitov R. Toll-like receptor control of the adaptive immune responses. Nat Immunol. 2004;5:987–995. - PubMed
-
- Weyrich AS, Zimmerman GA. Platelets: signaling cells in the immune continuum. Trends Immunol. 2004;25:489–495. - PubMed
-
- Joseph M. Immunopharmacology of Platelets. London, United Kingdom: Academic Press; 1995.
-
- Sharma A, Novak EK, Sojar HT, Swank RT, Kuramitsu HK, Genco RJ. Porphyromonas gingivalis platelet aggregation activity: outer membrane vesicles are potent activators of murine platelets. Oral Microbiol Immunol. 2000;15:393–396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
