

Development of monoclonal antibodies against human IRF-5 and their use in identifying the binding of IRF-5 to nuclear import proteins karyopherin-alpha1 and -beta1
- PMID: 19108028
- PMCID: PMC2628014
- DOI: 10.3349/ymj.2008.49.6.1023
Development of monoclonal antibodies against human IRF-5 and their use in identifying the binding of IRF-5 to nuclear import proteins karyopherin-alpha1 and -beta1
Abstract
Purpose: IRF-5 is a direct transducer of virus-mediated and TLR-mediated signaling pathways for the expression of cytokines and chemokines which form homodimers or heterodimers with IRF-7. However, direct IRF-5-specific monoclonal antibodies (mAbs) are not available at present. These could be used to further evaluate the functions of IRF-5. In this study, we produced and characterized three mouse mAbs to human IRF-5. The binding of IRF-5 to nuclear import proteins was first identified using a mAb.
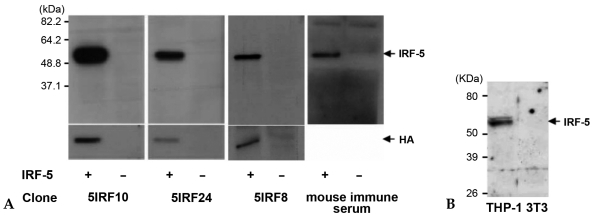
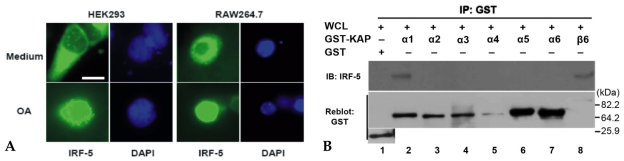
Materials and methods: His-tagged human IRF-5 protein spanning amino acid residues 193-257 was used as an antigen and three mAbs were produced. The mAbs were tested with ELISA, Western blot analysis (WB), immunofluorescent staining (IF), and immunoprecipitation (IP). In addition, the nuclear import protein which carried phosphorylated IRF-5 was identified using one of these mAbs.
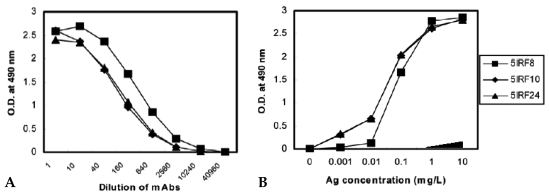
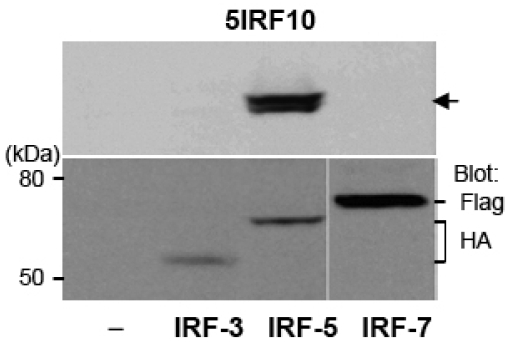
Results: MAbs 5IRF8, 5IRF10 and 5IRF24 which reacted with the recombinant His-IRF-5(193-257) protein were produced. All mAbs bound to human IRF-5, but not to IRF-3 or IRF-7. They could be used for WB, IF, and IP studies. The binding of phosphorylated IRF-5 to karyopherin-alpha1 and -beta1 was also identified.
Conclusion: Human IRF-5-specific mAbs are produced for studying the immunologic roles related to IRF-5. Phosphorylated IRF-5 is transported to the nucleus by binding to nuclear import proteins karyopherin-alpha1 and -beta1.
Figures



References
-
- Miyamoto M, Fujita T, Kimura Y, Maruyama M, Harada H, Sudo Y, et al. Regulated expression of a gene encoding a nuclear factor, IRF-1, that specifically binds to IFN-beta gene regulatory elements. Cell. 1988;54:903–913. - PubMed
-
- Lohoff M, Mak TW. Roles of interferon-regulatory factors in T-helper-cell differentiation. Nat Rev Immunol. 2005;5:125–135. - PubMed
-
- Taniguchi T, Ogasawara K, Takaoka A, Tanaka N. IRF family of transcription factors as regulators of host defense. Annu Rev Immunol. 2001;19:623–655. - PubMed
-
- Sato M, Suemori H, Hata N, Asagiri M, Ogasawara K, Nakao K, et al. Distinct and essential roles of transcription factors IRF-3 and IRF-7 in response to viruses for IFN-alpha/beta gene induction. Immunity. 2000;13:539–548. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
