

E2A acts in cis in G1 phase of cell cycle to promote Ig gene diversification
- PMID: 19109172
- PMCID: PMC2637363
- DOI: 10.4049/jimmunol.182.1.408
E2A acts in cis in G1 phase of cell cycle to promote Ig gene diversification
Abstract
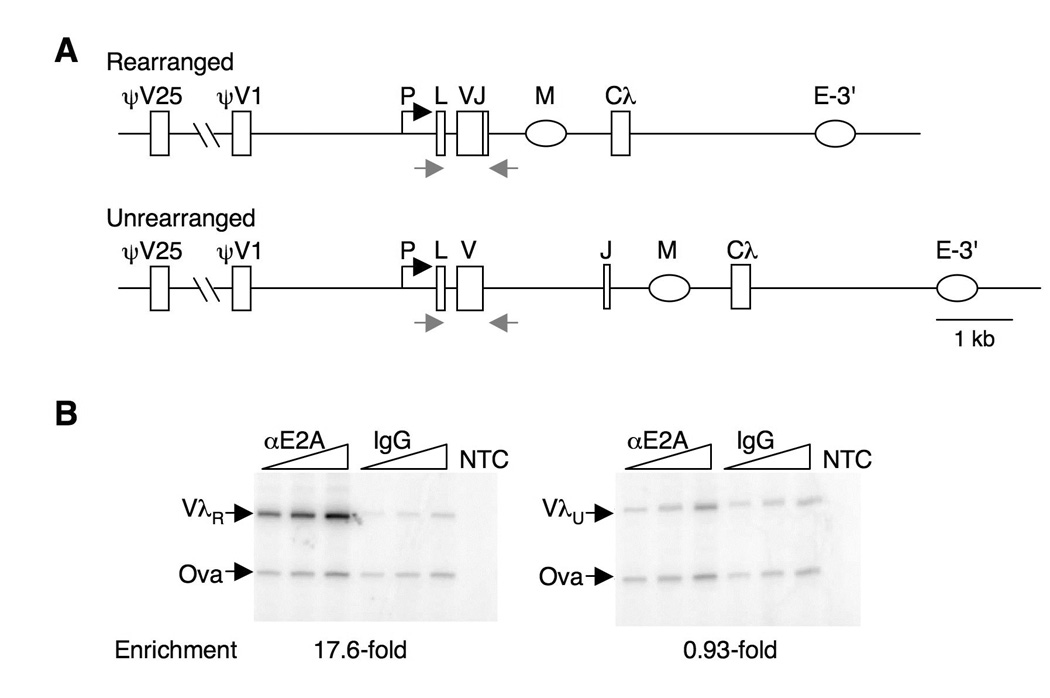
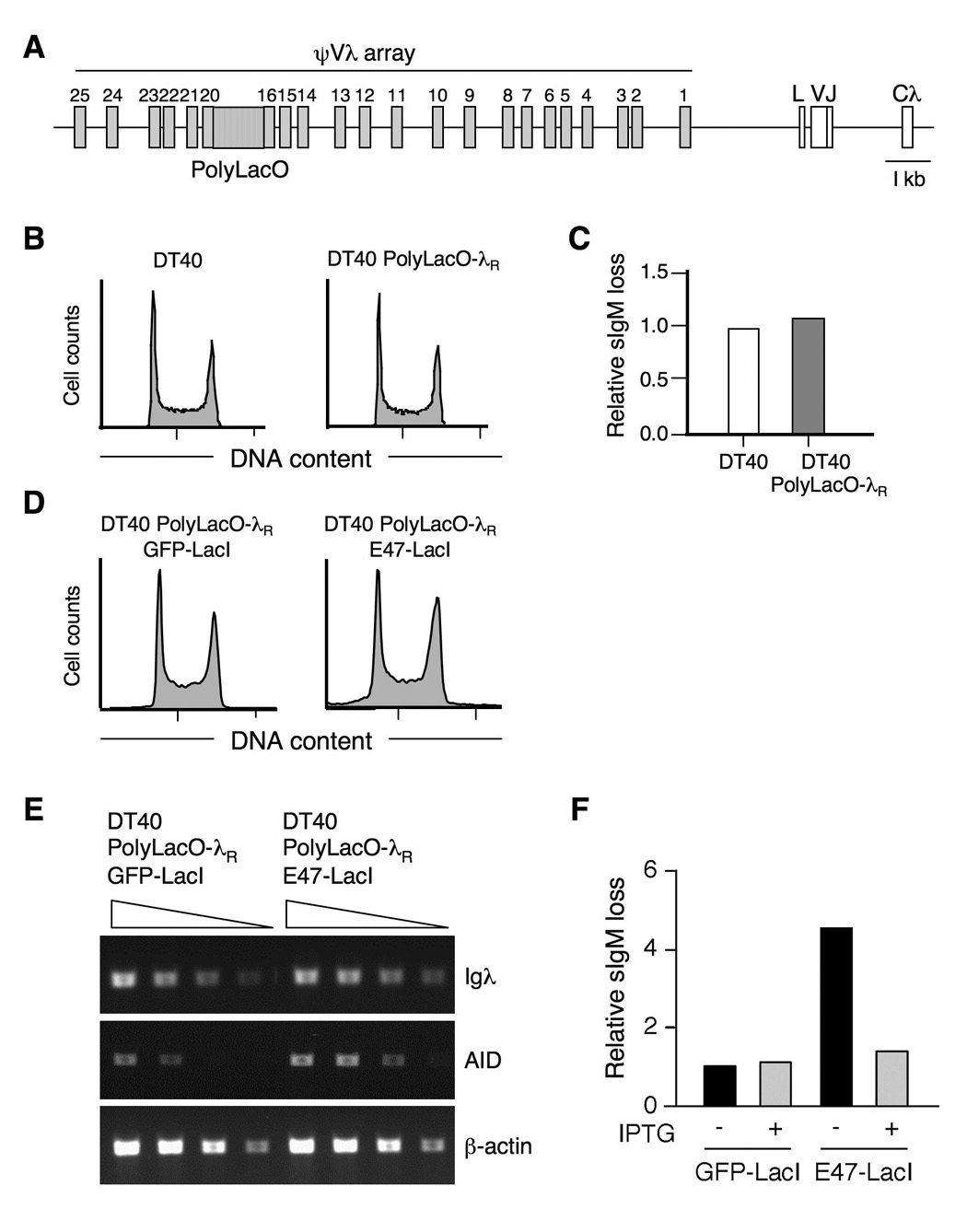
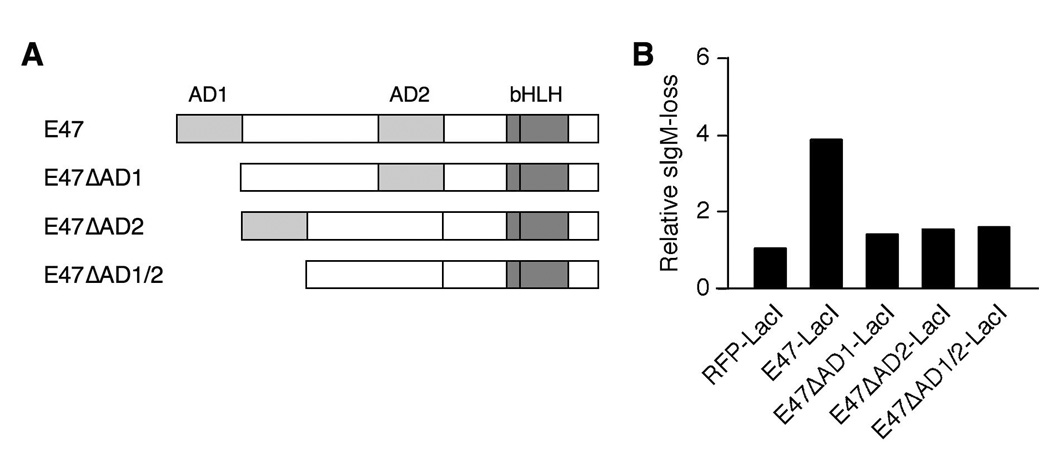
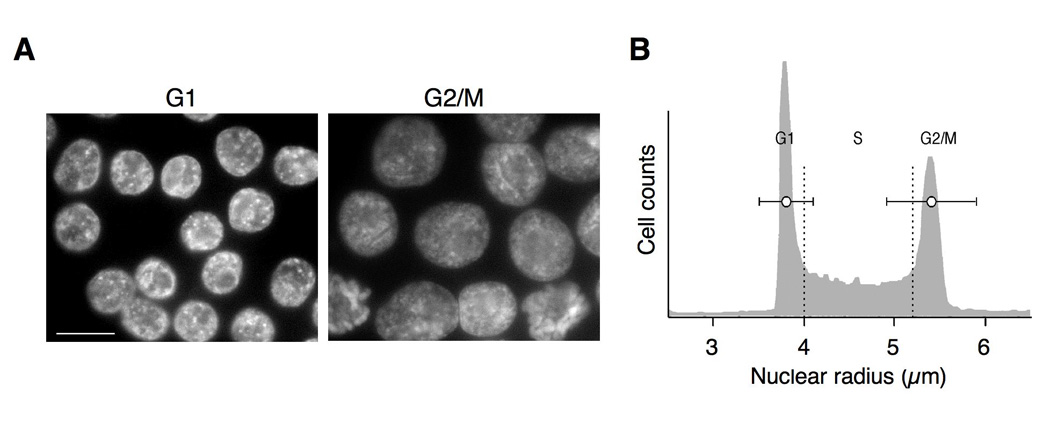
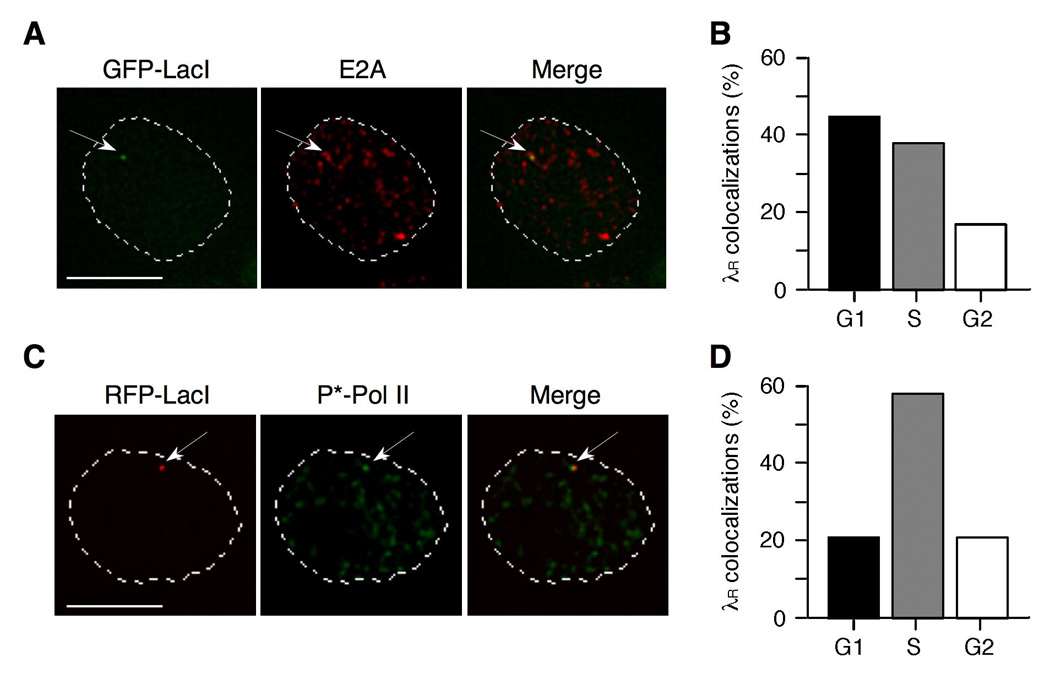
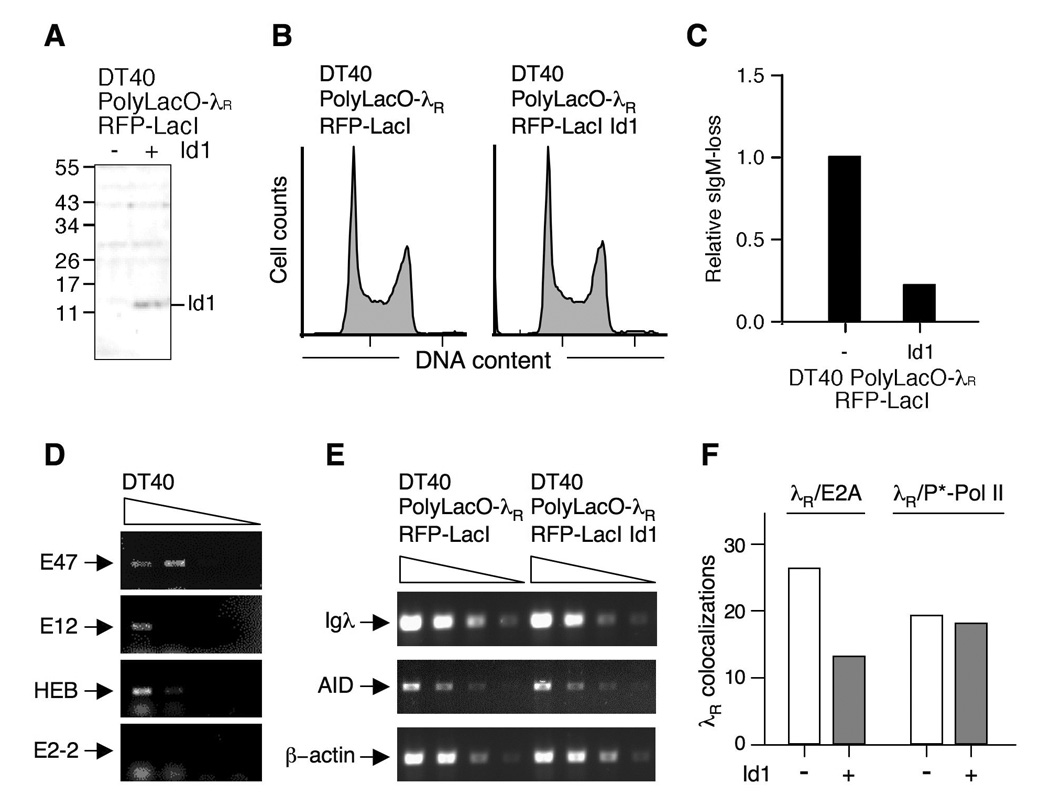
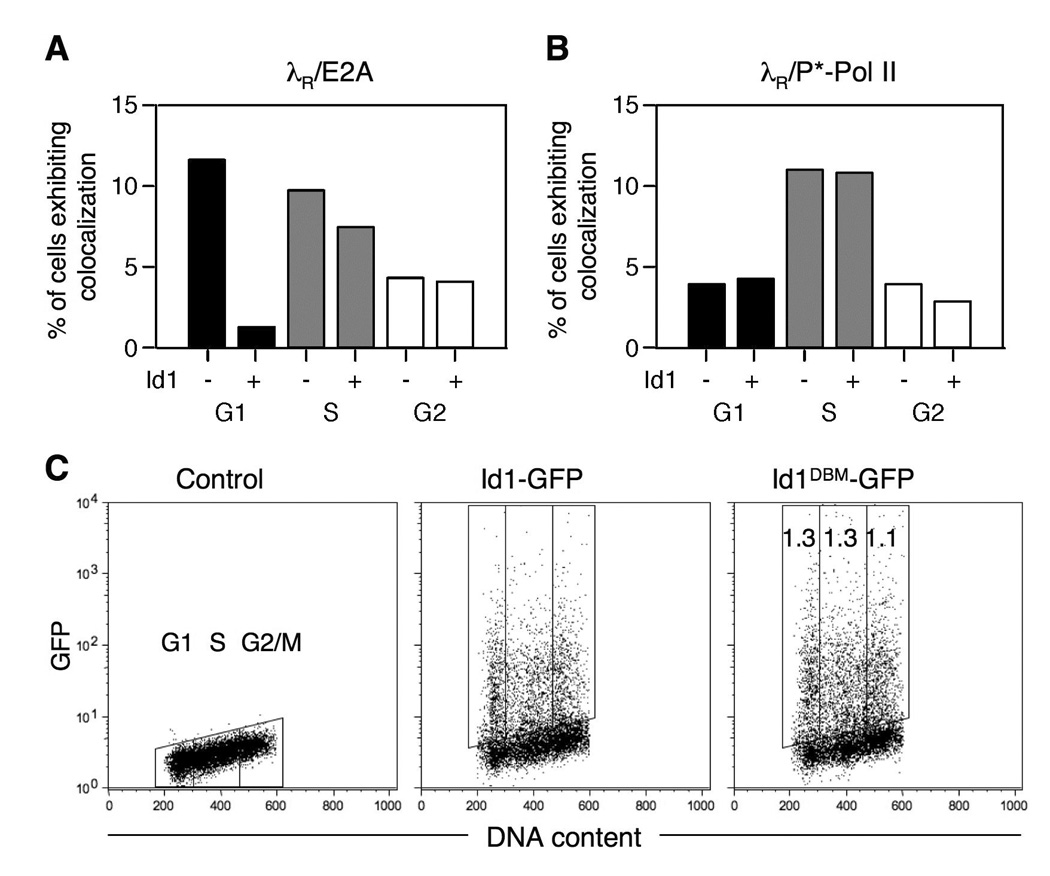
Rearranged Ig genes undergo diversification in sequence and structure initiated by the DNA deaminase, activation-induced deaminase. Ig genes must be transcribed for diversification to occur, but whether there are additional requirements for cis activation has not been established. Here we show, by chromatin immunoprecipitation, that the regulatory factor E2A associates with the rearranged Ig lambda(R) gene in the chicken DT40 B cell line, which performs constitutive Ig gene diversification. By analysis of a DT40 derivative in which polymerized lactose operator tags the rearranged lambda(R) gene, we show that E2A must function in cis to promote diversification and that stimulation of diversification in cis depends on the E2A activation domains. By direct imaging, we show that lambda(R)/E2A colocalizations are most prominent in G(1). We further show that expression of the E2A antagonist Id1 prevents lambda(R)/E2A colocalizations in G(1) and impairs diversification but not transcription of lambda(R). Thus, E2A acts in cis to promote Ig gene diversification, and G(1) phase is the critical window for E2A action.
Conflict of interest statement
The authors have no financial conflicts of interest.
Figures
Similar articles
-
Temporal regulation of Ig gene diversification revealed by single-cell imaging.J Immunol. 2009 Oct 1;183(7):4545-53. doi: 10.4049/jimmunol.0900673. Epub 2009 Sep 11. J Immunol. 2009. PMID: 19748985 Free PMC article.
-
Distinct roles for E12 and E47 in B cell specification and the sequential rearrangement of immunoglobulin light chain loci.J Exp Med. 2009 Sep 28;206(10):2271-84. doi: 10.1084/jem.20090756. Epub 2009 Sep 14. J Exp Med. 2009. PMID: 19752184 Free PMC article.
-
Regulation of histone H4 acetylation by transcription factor E2A in Ig gene conversion.Int Immunol. 2008 Feb;20(2):277-84. doi: 10.1093/intimm/dxm140. Epub 2008 Jan 7. Int Immunol. 2008. PMID: 18182382
-
Targeting of AID-mediated sequence diversification by cis-acting determinants.Adv Immunol. 2007;94:109-25. doi: 10.1016/S0065-2776(06)94004-8. Adv Immunol. 2007. PMID: 17560273 Review.
-
Gene rearrangement and B-cell development.Curr Opin Immunol. 1993 Apr;5(2):194-200. doi: 10.1016/0952-7915(93)90004-c. Curr Opin Immunol. 1993. PMID: 8507397 Review.
Cited by
-
Helix-Loop-Helix Proteins in Adaptive Immune Development.Front Immunol. 2022 May 12;13:881656. doi: 10.3389/fimmu.2022.881656. eCollection 2022. Front Immunol. 2022. PMID: 35634342 Free PMC article. Review.
-
Genetic diversification by somatic gene conversion.Genes (Basel). 2011 Jan 10;2(1):48-58. doi: 10.3390/genes2010048. Genes (Basel). 2011. PMID: 24710138 Free PMC article.
-
Temporal regulation of Ig gene diversification revealed by single-cell imaging.J Immunol. 2009 Oct 1;183(7):4545-53. doi: 10.4049/jimmunol.0900673. Epub 2009 Sep 11. J Immunol. 2009. PMID: 19748985 Free PMC article.
-
A critical context-dependent role for E boxes in the targeting of somatic hypermutation.J Immunol. 2013 Aug 15;191(4):1556-66. doi: 10.4049/jimmunol.1300969. Epub 2013 Jul 8. J Immunol. 2013. PMID: 23836058 Free PMC article.
-
RAD51 paralogs promote homology-directed repair at diversifying immunoglobulin V regions.BMC Mol Biol. 2009 Oct 28;10:98. doi: 10.1186/1471-2199-10-98. BMC Mol Biol. 2009. PMID: 19863810 Free PMC article.
References
-
- Maizels N. Immunoglobulin gene diversification. Annu. Rev. Genet. 2005;39:23–46. - PubMed
-
- Martomo SA, Gearhart PJ. Somatic hypermutation: subverted DNA repair. Curr. Opin. Immunol. 2006;18:243–248. - PubMed
-
- Di Noia JM, Neuberger MS. Molecular mechanisms of antibody somatic hypermutation. Annu. Rev. Biochem. 2007;76:1–22. - PubMed
-
- Teng G, Papavasiliou FN. Immunoglobulin somatic hypermutation. Annu. Rev. Genet. 2007;41:107–120. - PubMed
-
- Muramatsu M, Kinoshita K, Fagarasan S, Yamada S, Shinkai Y, Honjo T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 2000;102:553–563. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
