

Mice with a "monoclonal nose": perturbations in an olfactory map impair odor discrimination
- PMID: 19109912
- PMCID: PMC2732586
- DOI: 10.1016/j.neuron.2008.10.046
Mice with a "monoclonal nose": perturbations in an olfactory map impair odor discrimination
Abstract
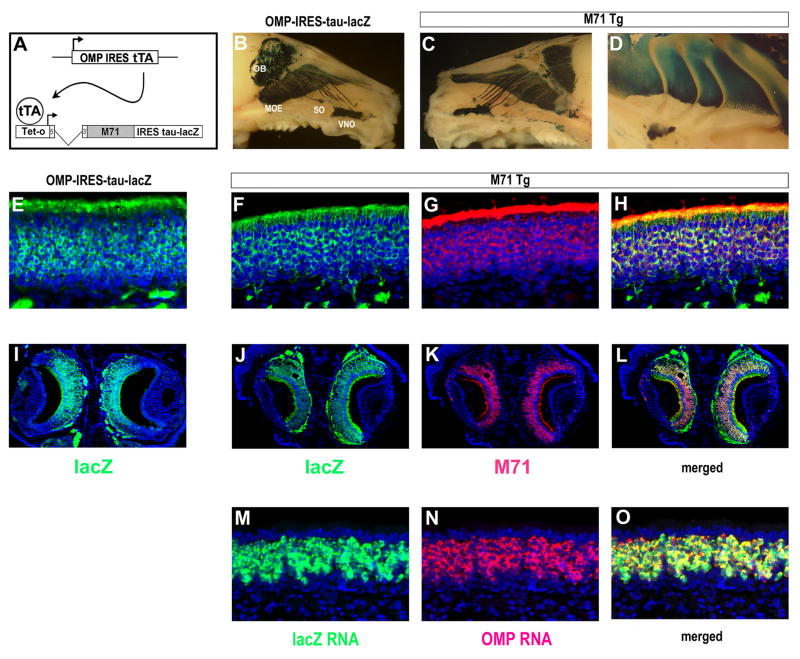
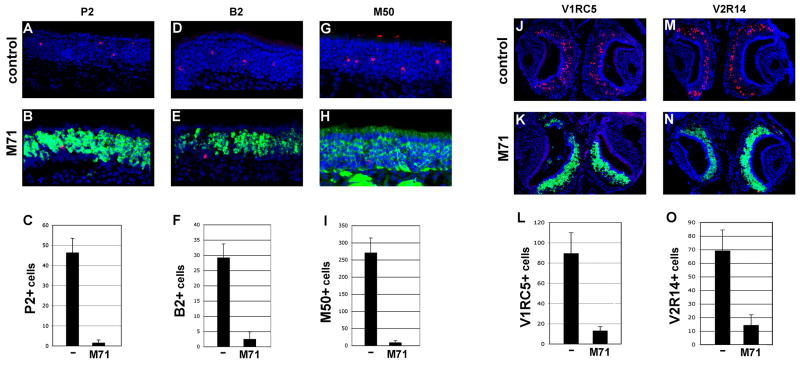
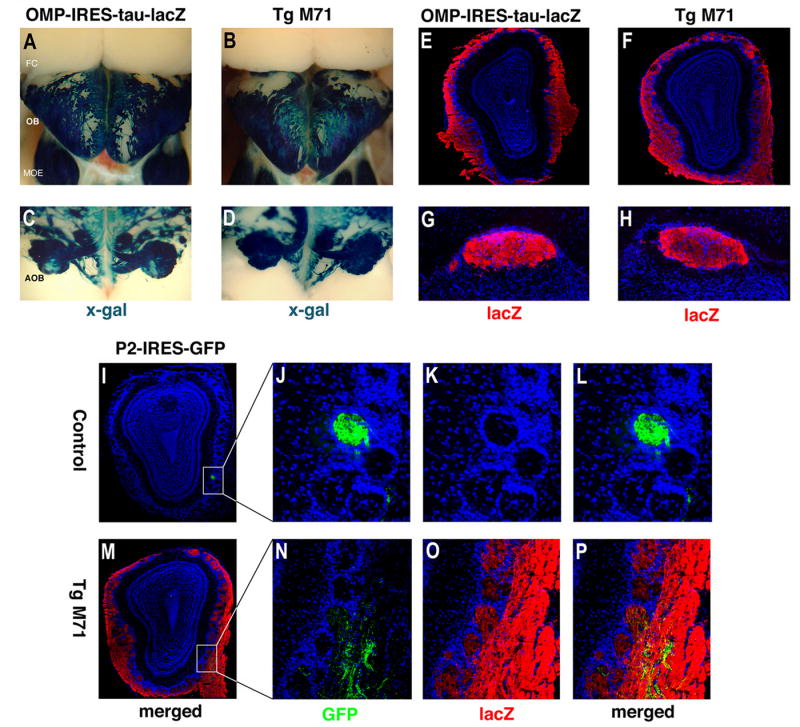
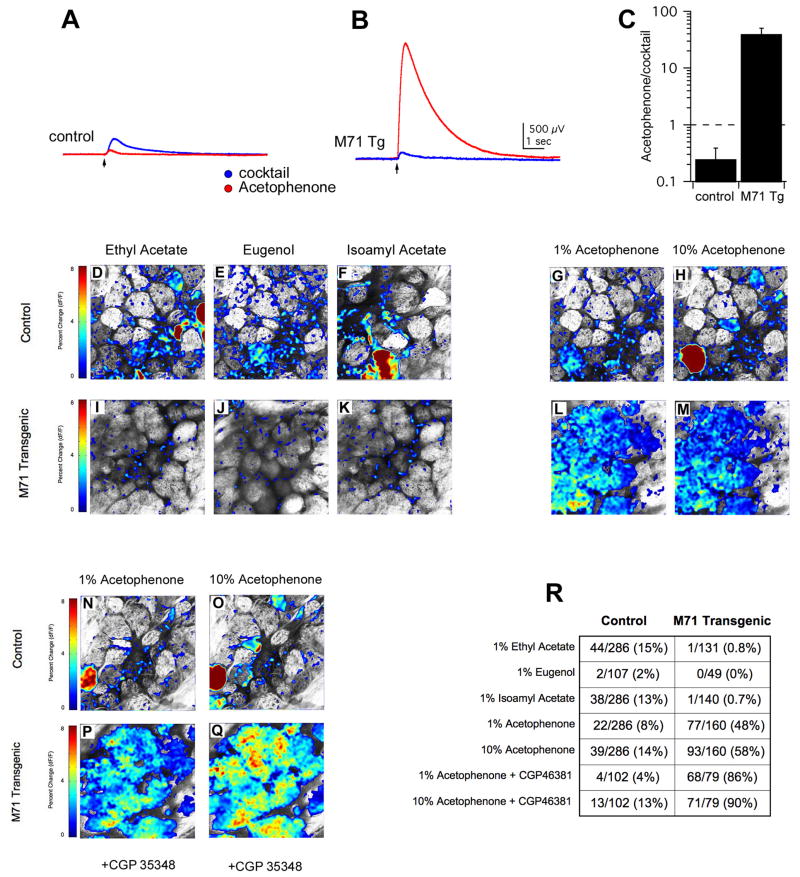
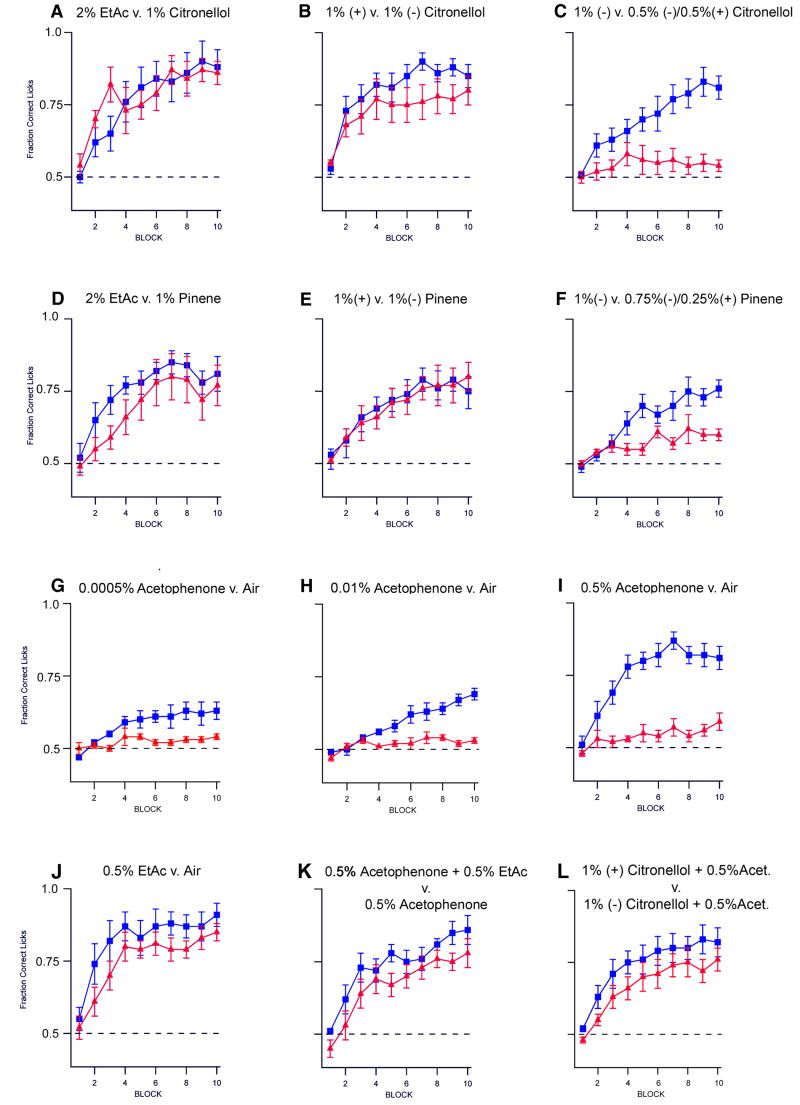
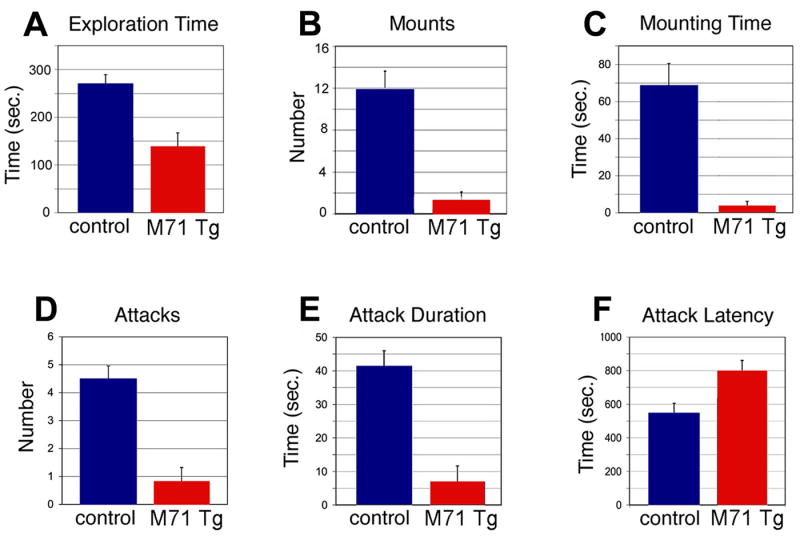
We have altered the neural representation of odors in the brain by generating a mouse with a "monoclonal nose" in which greater than 95% of the sensory neurons express a single odorant receptor, M71. As a consequence, the frequency of sensory neurons expressing endogenous receptor genes is reduced 20-fold. We observe that these mice can smell, but odor discrimination and performance in associative olfactory learning tasks are impaired. However, these mice cannot detect the M71 ligand acetophenone despite the observation that virtually all sensory neurons and glomeruli are activated by this odor. The M71 transgenic mice readily detect other odors in the presence of acetophenone. These observations have implications for how receptor activation in the periphery is represented in the brain and how these representations encode odors.
Figures
References
-
- Aroniadou-Anderjaska V, Zhou FM, Priest CA, Ennis M, Shipley MT. Tonic and synaptically evoked presynaptic inhibition of sensory input to the rat olfactory bulb via GABA(B) heteroreceptors. Journal of neurophysiology. 2000;84:1194–1203. - PubMed
-
- Aungst JL, Heyward PM, Puche AC, Karnup SV, Hayar A, Szabo G, Shipley MT. Centre-surround inhibition among olfactory bulb glomeruli. Nature. 2003;426:623–629. - PubMed
-
- Barlow HB. Single units and sensation: a neuron doctrine for perceptual psychology? Perception. 1972;1:371–394. - PubMed
-
- Bodyak N, Slotnick B. Performance of mice in an automated olfactometer: odor detection, discrimination and odor memory. Chemical senses. 1999;24:637–645. - PubMed
-
- Bozza T, McGann JP, Mombaerts P, Wachowiak M. In vivo imaging of neuronal activity by targeted expression of a genetically encoded probe in the mouse. Neuron. 2004;42:9–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
