

Probabilistic population codes for Bayesian decision making
- PMID: 19109917
- PMCID: PMC2742921
- DOI: 10.1016/j.neuron.2008.09.021
Probabilistic population codes for Bayesian decision making
Abstract
When making a decision, one must first accumulate evidence, often over time, and then select the appropriate action. Here, we present a neural model of decision making that can perform both evidence accumulation and action selection optimally. More specifically, we show that, given a Poisson-like distribution of spike counts, biological neural networks can accumulate evidence without loss of information through linear integration of neural activity and can select the most likely action through attractor dynamics. This holds for arbitrary correlations, any tuning curves, continuous and discrete variables, and sensory evidence whose reliability varies over time. Our model predicts that the neurons in the lateral intraparietal cortex involved in evidence accumulation encode, on every trial, a probability distribution which predicts the animal's performance. We present experimental evidence consistent with this prediction and discuss other predictions applicable to more general settings.
Figures

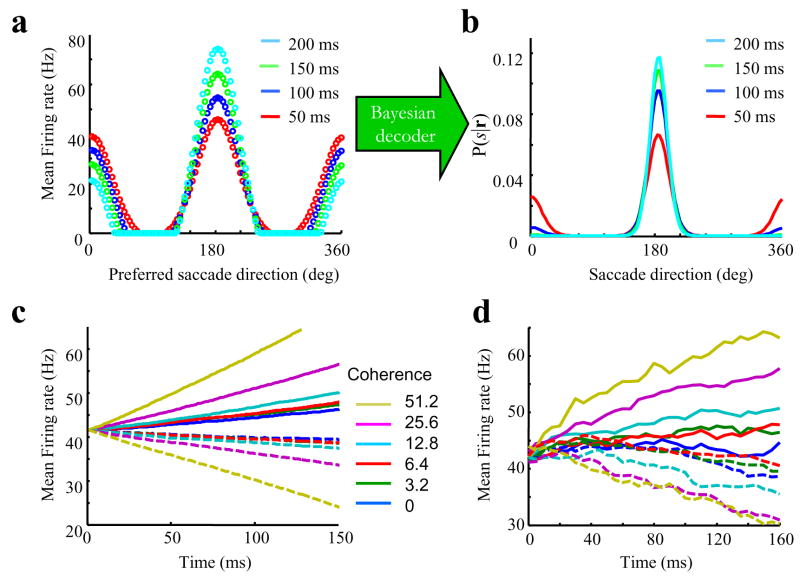

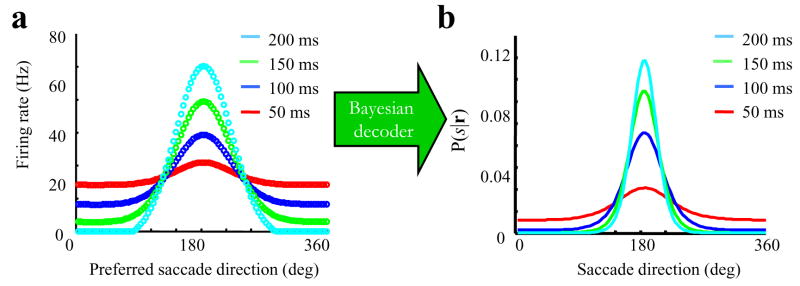


Comment in
-
So many choices: what computational models reveal about decision-making mechanisms.Neuron. 2008 Dec 26;60(6):946-9. doi: 10.1016/j.neuron.2008.12.011. Neuron. 2008. PMID: 19109902
References
-
- Anderson JS, Lampl I, Gillespie DC, Ferster D. The contribution of noise to contrast invariance of orientation tuning in cat visual cortex. Science. 2000;290:1968–1972. - PubMed
-
- Barber MJ, Clark JW, Anderson CH. Neural representation of probabilistic information. Neural Comput. 2003;15:1843–1864. - PubMed
-
- Bogacz R, Brown E, Moehlis J, Holmes P, Cohen JD. The physics of optimal decision making: a formal analysis of models of performance in two-alternative forced-choice tasks. Psychol Rev. 2006;113:700–765. - PubMed
-
- Bogacz R, Gurney K. The basal ganglia and cortex implement optimal decision making between alternative actions. Neural Comput. 2007;19:442–477. - PubMed
-
- Britten KH, Shadlen MN, Newsome WT, Movshon JA. Responses of neurons in macaque MT to stochastic motion signals. Vis Neurosci. 1993;10:1157–1169. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
