

Similarity effect and optimal control of multiple-choice decision making
- PMID: 19109918
- PMCID: PMC2633638
- DOI: 10.1016/j.neuron.2008.12.003
Similarity effect and optimal control of multiple-choice decision making
Abstract
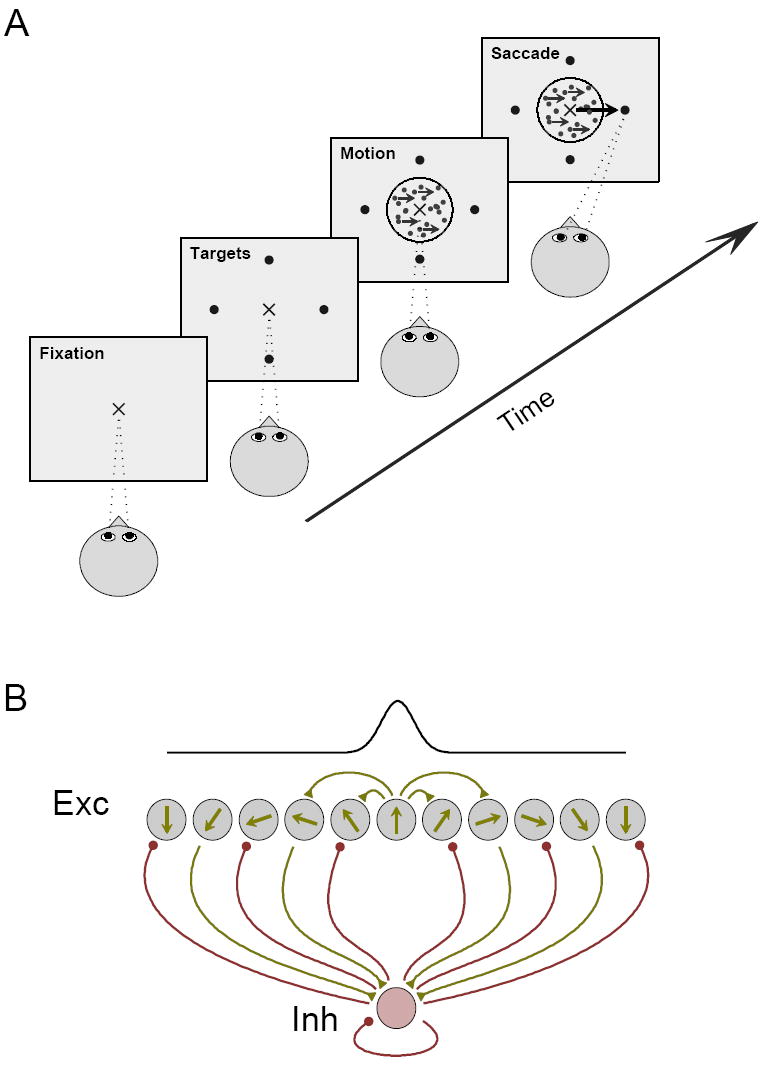
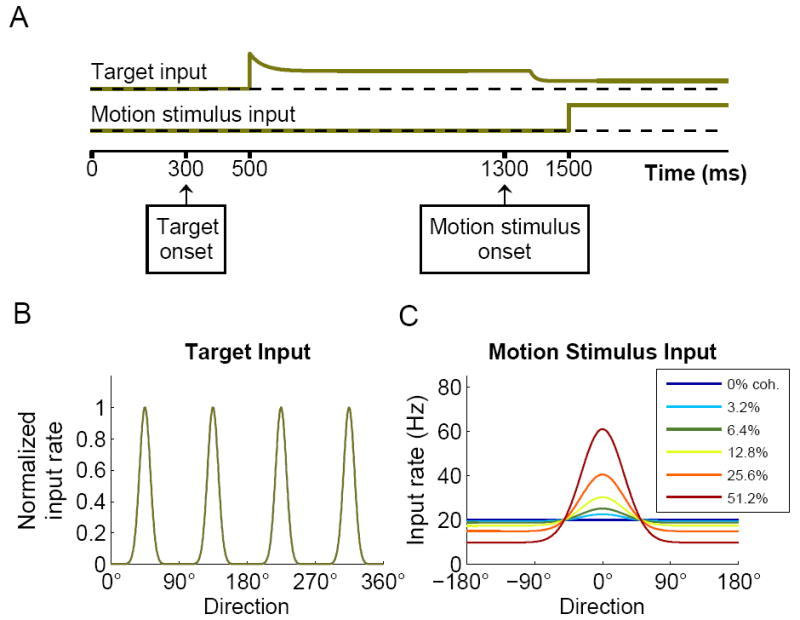
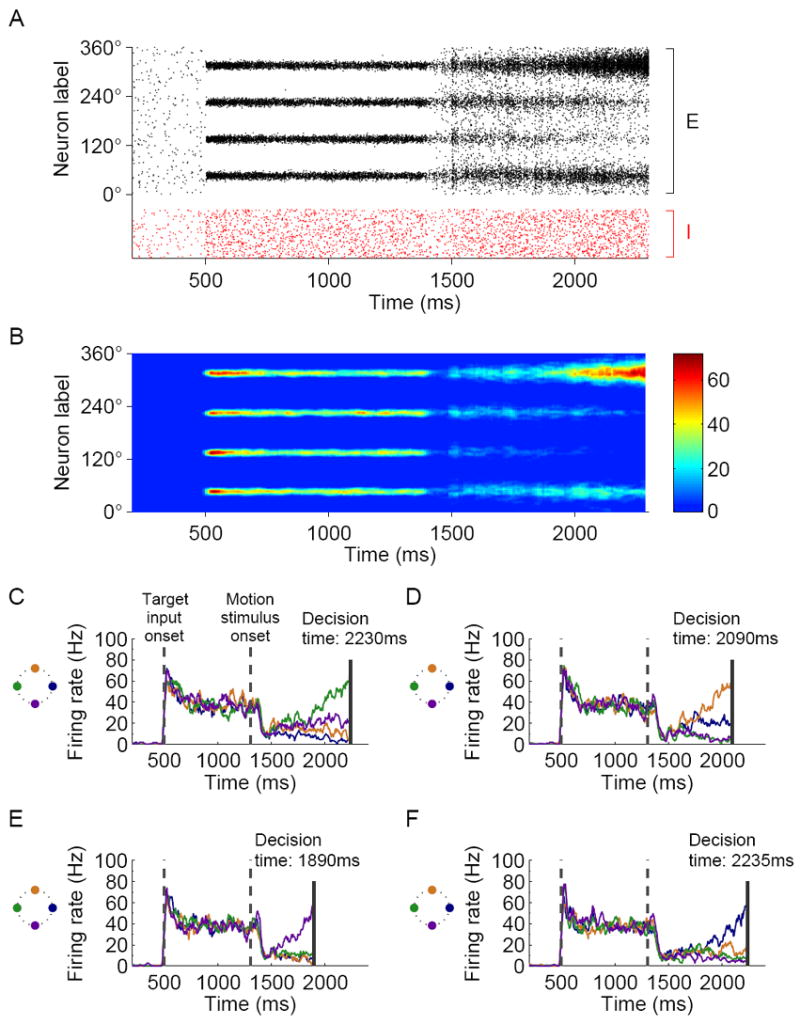
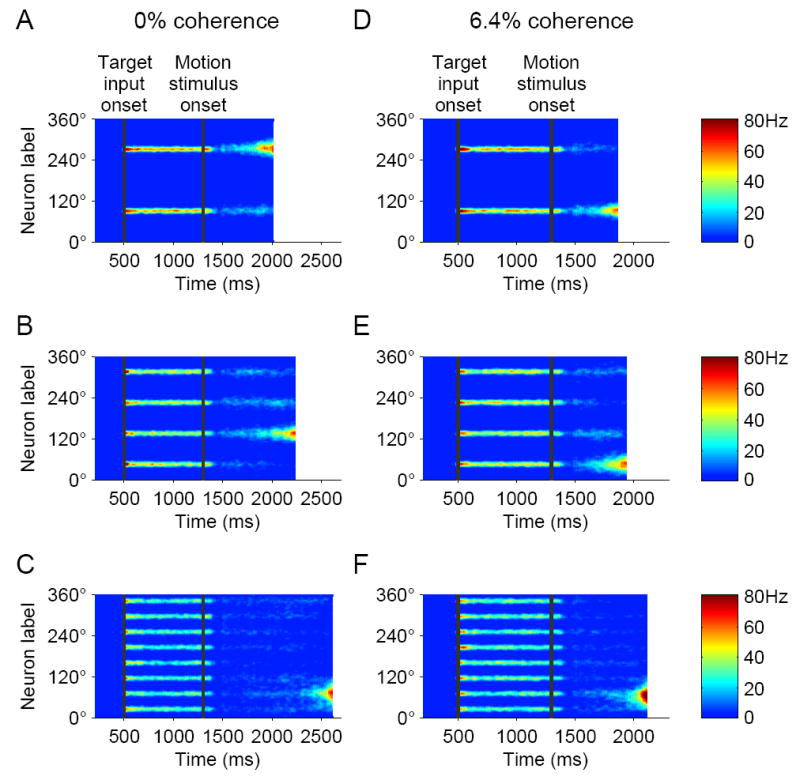
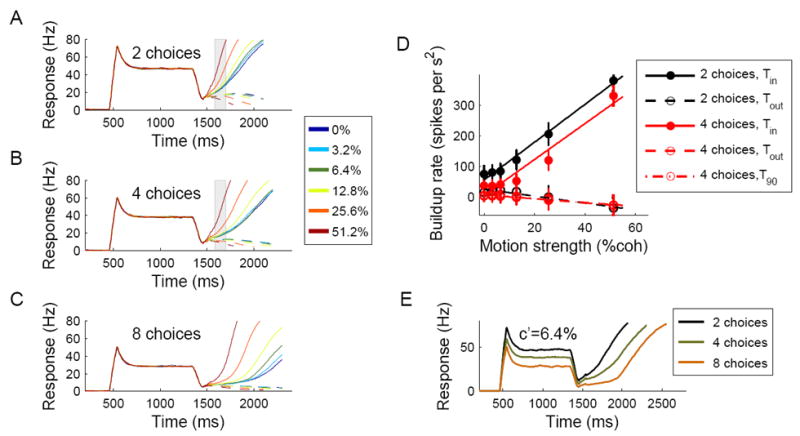
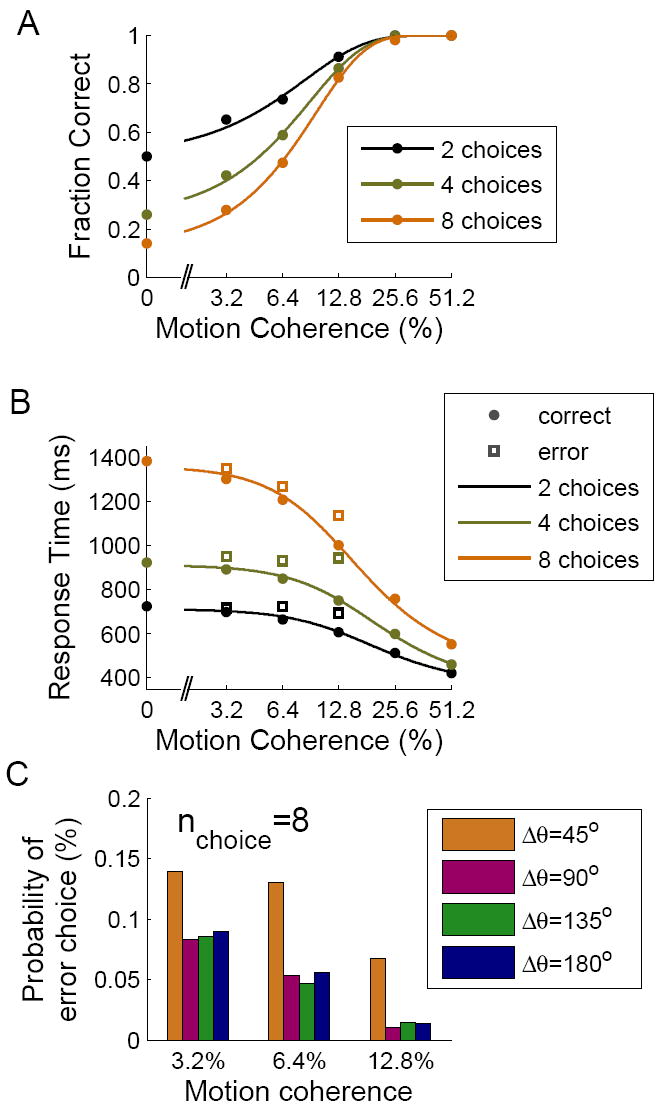
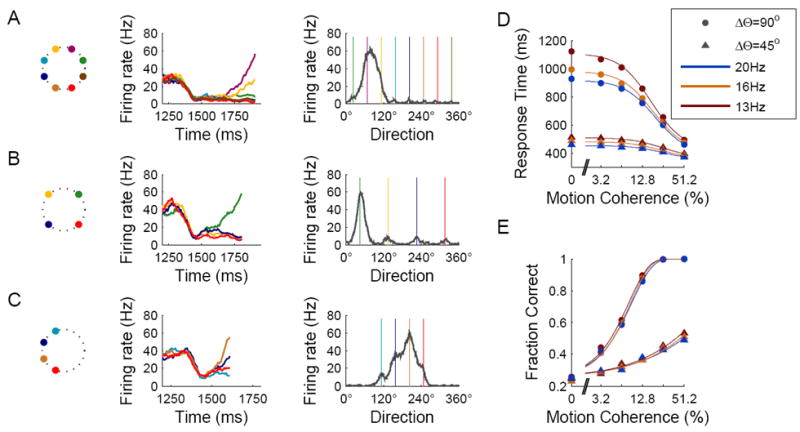
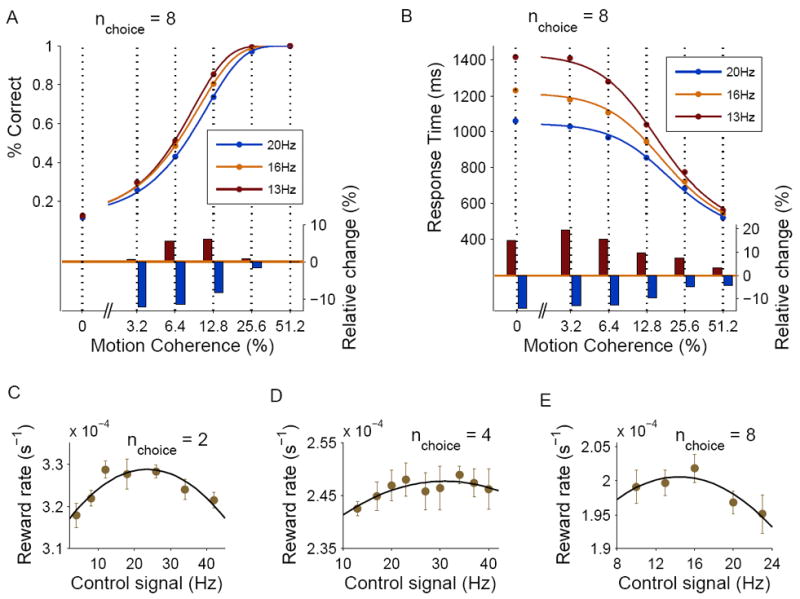
Decision making with several choice options is central to cognition. To elucidate the neural mechanisms of such decisions, we investigated a recurrent cortical circuit model in which fluctuating spiking neural dynamics underlie trial-by-trial stochastic decisions. The model encodes a continuous analog stimulus feature and is thus applicable to multiple-choice decisions. Importantly, the continuous network captures similarity between alternatives and possible overlaps in their neural representation. Model simulations accounted for behavioral as well as single-unit neurophysiological data from a recent monkey experiment and revealed testable predictions about the patterns of error rate as a function of the similarity between the correct and actual choices. We also found that the similarity and number of options affect speed and accuracy of responses. A mechanism is proposed for flexible control of speed-accuracy tradeoff, based on a simple top-down signal to the decision circuit that may vary nonmonotonically with the number of choice alternatives.
Figures
Comment in
-
So many choices: what computational models reveal about decision-making mechanisms.Neuron. 2008 Dec 26;60(6):946-9. doi: 10.1016/j.neuron.2008.12.011. Neuron. 2008. PMID: 19109902
References
-
- Ashby FG, Perrin NA. Toward a unified theory of similarity and recognition. Psychological Review. 1988;95:124–150.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
