

Design and signaling mechanism of light-regulated histidine kinases
- PMID: 19109976
- PMCID: PMC3527124
- DOI: 10.1016/j.jmb.2008.12.017
Design and signaling mechanism of light-regulated histidine kinases
Abstract
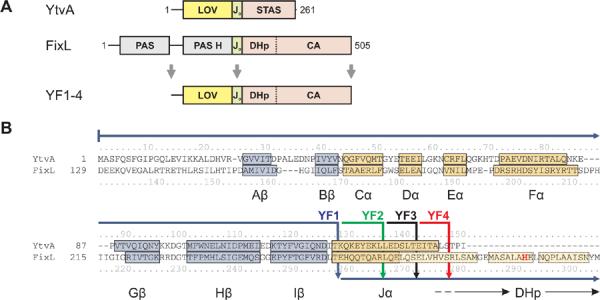
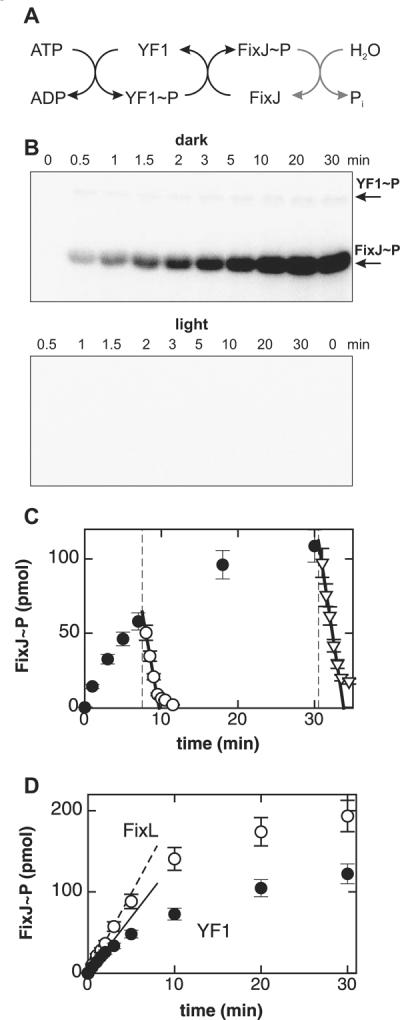
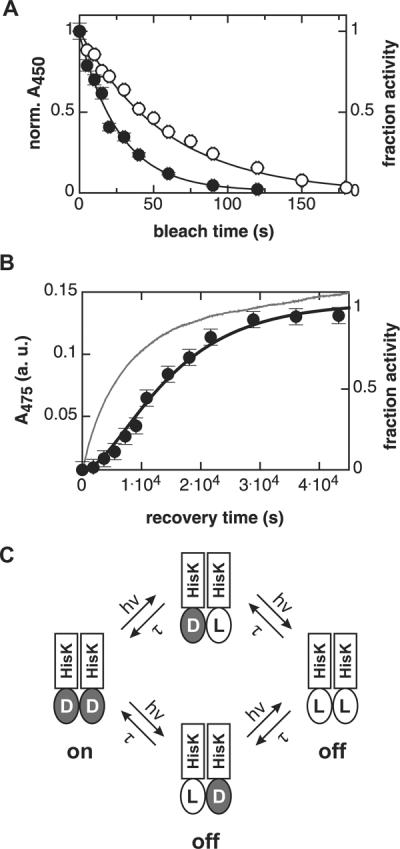
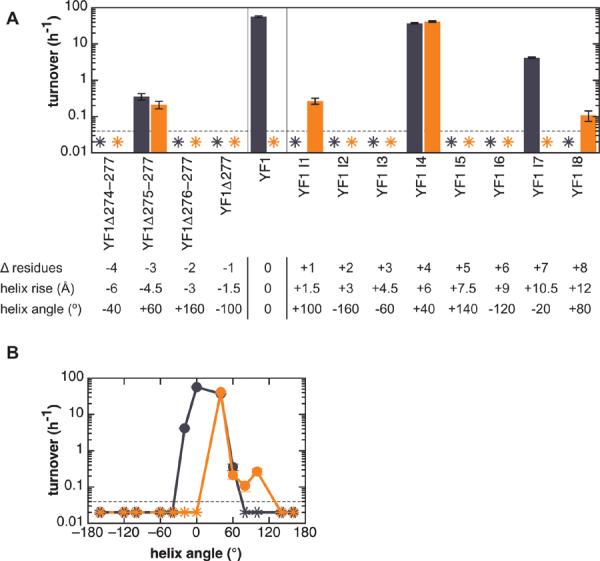
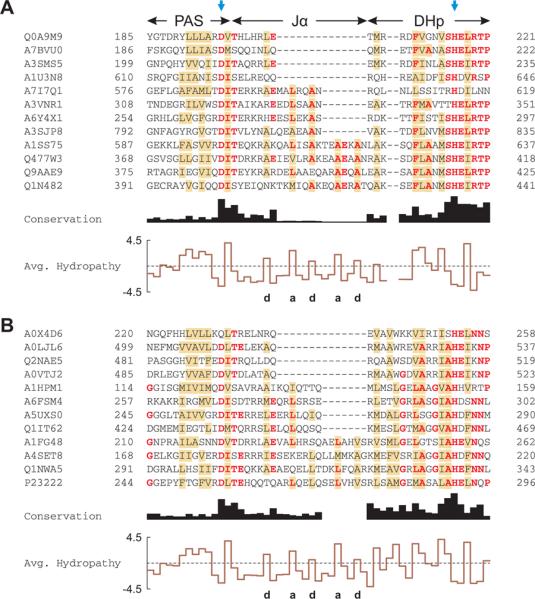
Signal transduction proteins are organized into sensor (input) domains that perceive a signal and, in response, regulate the biological activity of effector (output) domains. We reprogrammed the input signal specificity of a normally oxygen-sensitive, light-inert histidine kinase by replacing its chemosensor domain by a light-oxygen-voltage photosensor domain. Illumination of the resultant fusion kinase YF1 reduced net kinase activity by approximately 1000-fold in vitro. YF1 also controls gene expression in a light-dependent manner in vivo. Signals are transmitted from the light-oxygen-voltage sensor domain to the histidine kinase domain via a 40 degrees -60 degrees rotational movement within an alpha-helical coiled-coil linker; light is acting as a rotary switch. These signaling principles are broadly applicable to domains linked by alpha-helices and to chemo- and photosensors. Conserved sequence motifs guide the rational design of light-regulated variants of histidine kinases and other proteins.
Figures
References
-
- Pawson T, Nash P. Assembly of cell regulatory systems through protein interaction domains. Science. 2003;300:445–52. - PubMed
-
- Dueber JE, Yeh BJ, Chak K, Lim WA. Reprogramming control of an allosteric signaling switch through modular recombination. Science. 2003;301:1904–8. - PubMed
-
- Bashor CJ, Helman NC, Yan S, Lim WA. Using engineered scaffold interactions to reshape MAP kinase pathway signaling dynamics. Science. 2008;319:1539–43. - PubMed
-
- Christie JM, Reymond P, Powell GK, Bernasconi P, Raibekas AA, Liscum E, Briggs WR. Arabidopsis NPH1: a flavoprotein with the properties of a photoreceptor for phototropism. Science. 1998;282:1698–701. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
