

Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to form the premetastatic niche
- PMID: 19111879
- PMCID: PMC3050620
- DOI: 10.1016/j.ccr.2008.11.012
Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to form the premetastatic niche
Abstract
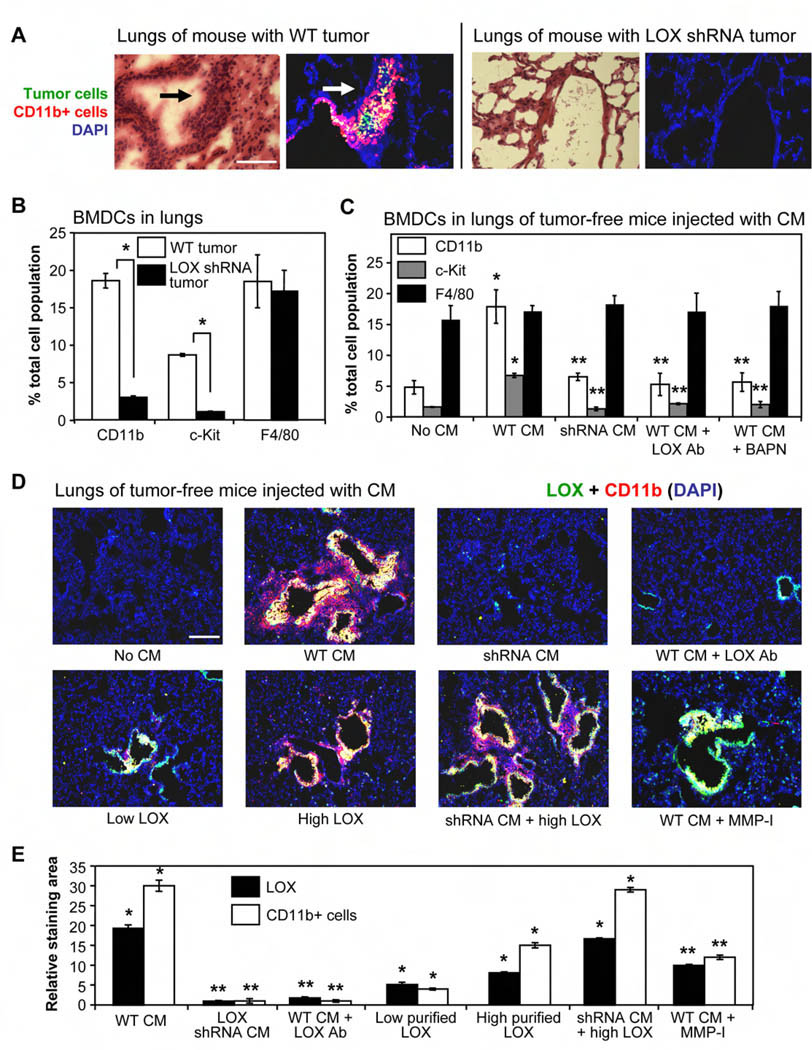
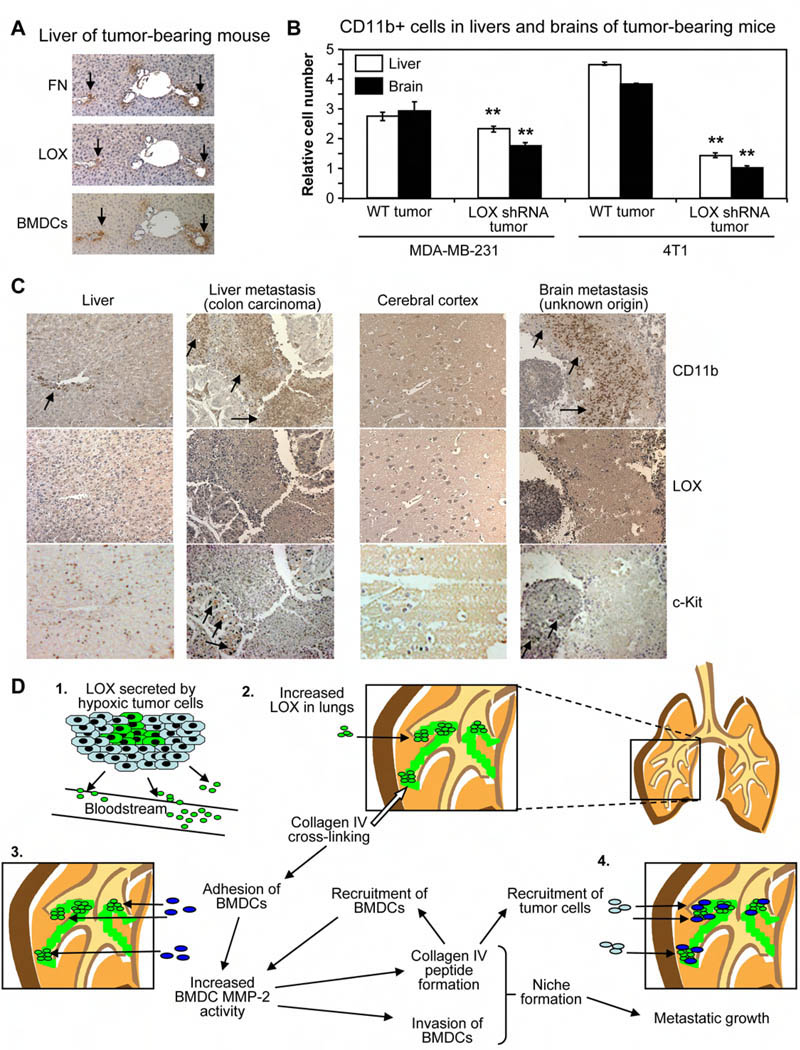
Tumor cell metastasis is facilitated by "premetastatic niches" formed in destination organs by invading bone marrow-derived cells (BMDCs). Lysyl oxidase (LOX) is critical for premetastatic niche formation. LOX secreted by hypoxic breast tumor cells accumulates at premetastatic sites, crosslinks collagen IV in the basement membrane, and is essential for CD11b+ myeloid cell recruitment. CD11b+ cells adhere to crosslinked collagen IV and produce matrix metalloproteinase-2, which cleaves collagen, enhancing the invasion and recruitment of BMDCs and metastasizing tumor cells. LOX inhibition prevents CD11b+ cell recruitment and metastatic growth. CD11b+ cells and LOX also colocalize in biopsies of human metastases. Our findings demonstrate a critical role for LOX in premetastatic niche formation and support targeting LOX for the treatment and prevention of metastatic disease.
Figures


References
-
- Cairns RA, Khokha R, Hill RP. Molecular mechanisms of tumor invasion and metastasis: an integrated view. Curr Mol Med. 2003;3:659–671. - PubMed
-
- Cameron JD, Skubitz AP, Furcht LT. Type IV collagen and corneal epithelial adhesion and migration. Effects of type IV collagen fragments and synthetic peptides on rabbit corneal epithelial cell adhesion and migration in vitro. Invest Ophthalmol Vis Sci. 1991;32:2766–2773. - PubMed
-
- Condeelis J, Pollard JW. Macrophages: obligate partners for tumor cell migration, invasion, and metastasis. Cell. 2006;124:263–266. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
