

Pivotal Advance: Toll-like receptor regulation of scavenger receptor-A-mediated phagocytosis
- PMID: 19112093
- PMCID: PMC2656429
- DOI: 10.1189/jlb.1008631
Pivotal Advance: Toll-like receptor regulation of scavenger receptor-A-mediated phagocytosis
Abstract
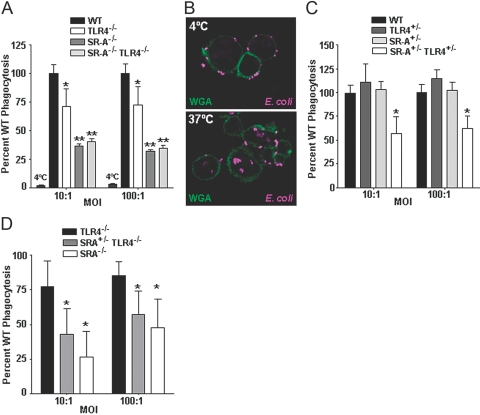
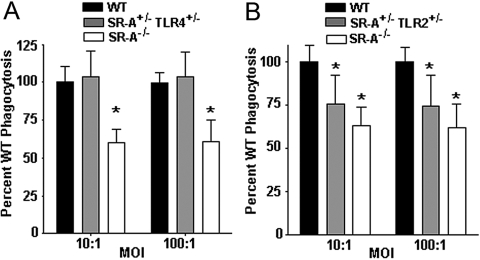
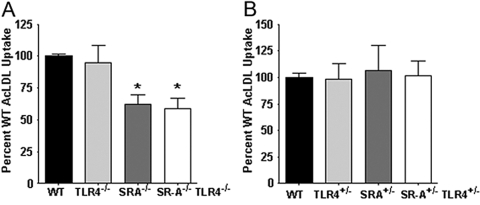
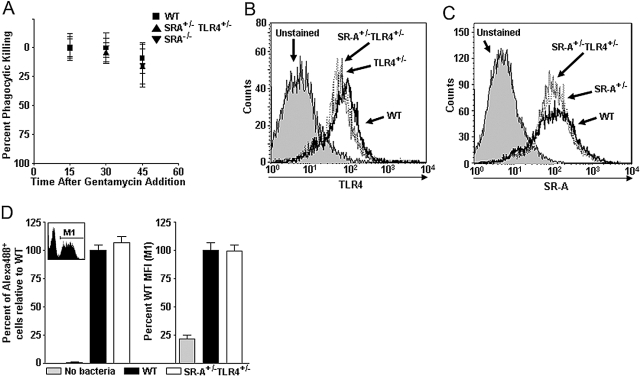
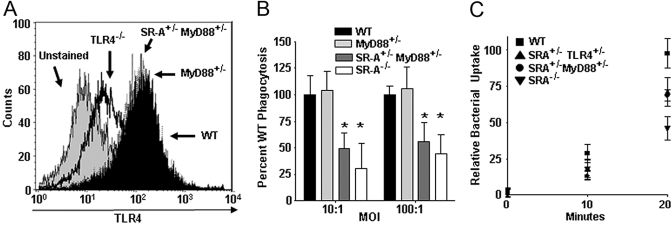
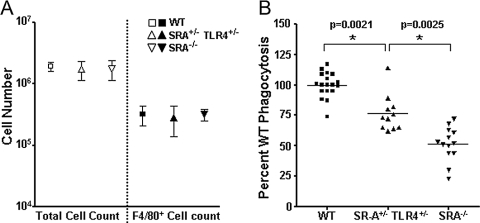
Class-A scavenger receptors (SR-A) and TLR mediate early immune responses against pathogenic bacteria. SR-A and TLR molecules are expressed on phagocytes and interact with common ligands from Gram-negative and Gram-positive bacteria; however, the contribution of TLR activity to SR-A-mediated phagocytosis has not been assessed directly. Herein, we provide genetic and functional evidence that ligand- and TLR-specific stimuli synergize with SR-A to mediate bacterial phagocytosis. Although complete loss of SR-A (SR-A(-/-)) is known to impair bacterial clearance, here we identify the first deficiency attributable to SR-A heterozygosity: SR-A(+/-)TLR4(+/-) cells and mice are impaired significantly in the clearance of Gram-negative Escherichia coli. This phenotype is specific to the TLR signaling event, as SR-A(+/-)TLR4(+/-) cells are not deficient for the clearance of Gram-positive Staphylococcus aureus bacteria, which contain cell-surface TLR2 ligands but lack TLR4 ligands. We demonstrate that this is a global, phagocytic mechanism, regulated independently by multiple TLRs, as analogous to the SR-A(+/-)TLR4(+/-) deficit, SR-A(+/-)TLR2(+/-) cells are impaired for S. aureus uptake. In support of this, we show that SR-A(+/-)MyD88(+/-) cells recapitulate the phagocytosis defect observed in SR-A(+/-)TLR4(+/-) cells. These data identify for the first time that TLR-driven innate immune responses, via a MyD88 signaling mechanism, regulate SR-A-dependent phagocytosis of bacteria. These findings provide novel insights into how innate immune cells control SR-A-mediated trafficking and are the first demonstration that subtle changes in the expression of SR-A and TLRs can substantially affect host bacterial clearance.
Figures
References
-
- Janeway C A, Jr, Medzhitov R. Innate immune recognition. Annu Rev Immunol. 2002;20:197–216. - PubMed
-
- Medzhitov R, Janeway C A., Jr Decoding the patterns of self and nonself by the innate immune system. Science. 2002;296:298–300. - PubMed
-
- Blander J M, Medzhitov R. Regulation of phagosome maturation by signals from Toll-like receptors. Science. 2004;304:1014–1018. - PubMed
-
- Blander J M, Medzhitov R. On regulation of phagosome maturation and antigen presentation. Nat Immunol. 2006;7:1029–1035. - PubMed
-
- Gough P J, Gordon S. The role of scavenger receptors in the innate immune system. Microbes Infect. 2000;2:305–311. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
