

Characterization of a Coxiella burnetii ftsZ mutant generated by Himar1 transposon mutagenesis
- PMID: 19114492
- PMCID: PMC2648191
- DOI: 10.1128/JB.01580-08
Characterization of a Coxiella burnetii ftsZ mutant generated by Himar1 transposon mutagenesis
Abstract
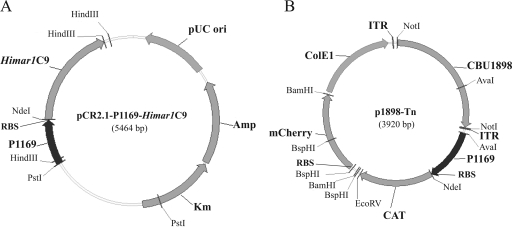
Coxiella burnetii is a gram-negative obligate intracellular bacterium and the causative agent of human Q fever. The lack of methods to genetically manipulate C. burnetii significantly impedes the study of this organism. We describe here the cloning and characterization of a C. burnetii ftsZ mutant generated by mariner-based Himar1 transposon (Tn) mutagenesis. C. burnetii was coelectroporated with a plasmid encoding the Himar1 C9 transposase variant and a plasmid containing a Himar1 transposon encoding chloramphenicol acetyltransferase, mCherry fluorescent protein, and a ColE1 origin of replication. Vero cells were infected with electroporated C. burnetii and transformants scored as organisms replicating in the presence of chloramphenicol and expressing mCherry. Southern blot analysis revealed multiple transpositions in the C. burnetii genome and rescue cloning identified 30 and 5 insertions in coding and noncoding regions, respectively. Using micromanipulation, a C. burnetii clone was isolated containing a Tn insertion within the C terminus of the cell division gene ftsZ. The ftsZ mutant had a significantly lower growth rate than wild-type bacteria and frequently appeared as filamentous forms displaying incomplete cell division septa. The latter phenotype correlated with a deficiency in generating infectious foci on a per-genome basis compared to wild-type organisms. The mutant FtsZ protein was also unable to bind the essential cell division protein FtsA. This is the first description of C. burnetii harboring a defined gene mutation generated by genetic transformation.
Figures
References
-
- Chen, S. Y., T. A. Hoover, H. A. Thompson, and J. C. Williams. 1990. Characterization of the origin of DNA replication of the Coxiella burnetii chromosome. Ann. N. Y. Acad. Sci. 590491-503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
