

Hypovirus-responsive transcription factor gene pro1 of the chestnut blight fungus Cryphonectria parasitica is required for female fertility, asexual spore development, and stable maintenance of hypovirus infection
- PMID: 19114501
- PMCID: PMC2653244
- DOI: 10.1128/EC.00338-08
Hypovirus-responsive transcription factor gene pro1 of the chestnut blight fungus Cryphonectria parasitica is required for female fertility, asexual spore development, and stable maintenance of hypovirus infection
Abstract
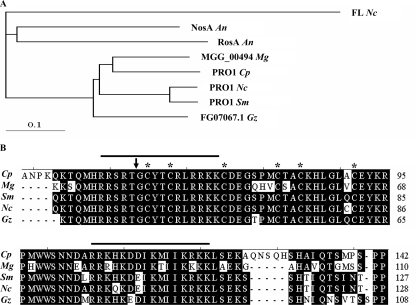
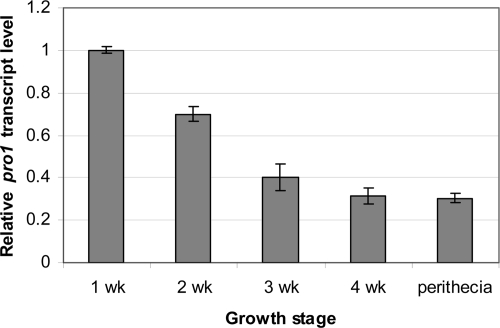
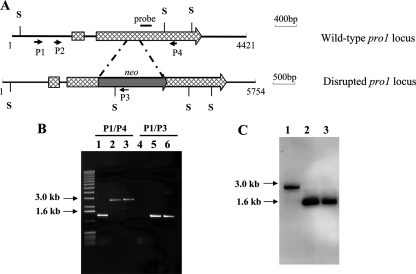
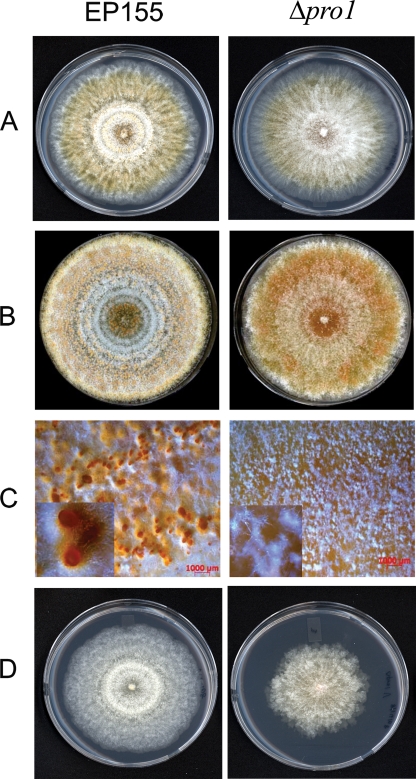
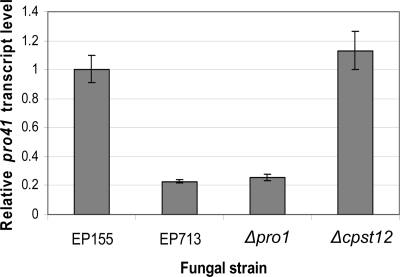
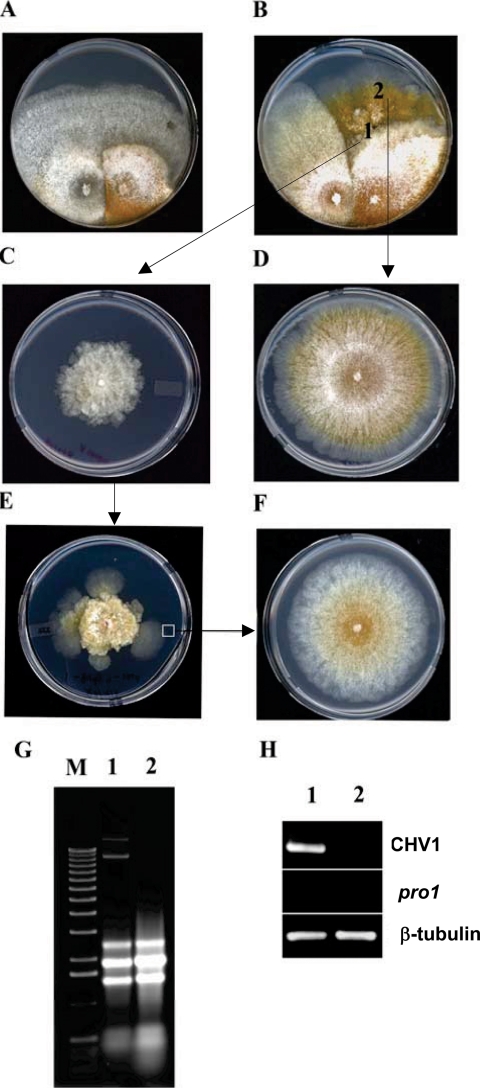
We report characterization of the gene encoding putative transcription factor PRO1, identified in transcriptional profiling studies as being downregulated in the chestnut blight fungus Cryphonectria parasitica in response to infection by virulence-attenuating hypoviruses. Sequence analysis confirmed that pro1 encodes a Zn(II)(2)Cys(6) binuclear cluster DNA binding protein with significant sequence similarity to the pro1 gene product that controls fruiting body development in Sordaria macrospora. Targeted disruption of the C. parasitica pro1 gene resulted in two phenotypic changes that also accompany hypovirus infection, a significant reduction in asexual sporulation that could be reversed by exposure to high light intensity, and loss of female fertility. The pro1 disruption mutant, however, retained full virulence. Although hypovirus CHV1-EP713 infection was established in the pro1 disruption mutant, infected colonies continually produced virus-free sectors, suggesting that PRO1 is required for stable maintenance of hypovirus infection. These results complement the recent characterization of the hypovirus-responsive homologue of the Saccharomyces cerevisiae Ste12 C(2)H(2) zinc finger transcription factor gene, cpst12, which was shown to be required for C. parasitica female fertility and virulence.
Figures

References
-
- Anagnostakis, S. L. 1982. Biological control of chestnut blight. Science 215466-471. - PubMed
-
- Anagnostakis, S. L. 1984. Nuclear gene mutations in Endothia (Cryphonectria) parasitica that affect morphology and virulence. Phytopathology 74761-765.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
