

Coordinated control of self-renewal and differentiation of neural stem cells by Myc and the p19ARF-p53 pathway
- PMID: 19114593
- PMCID: PMC2606961
- DOI: 10.1083/jcb.200807130
Coordinated control of self-renewal and differentiation of neural stem cells by Myc and the p19ARF-p53 pathway
Abstract
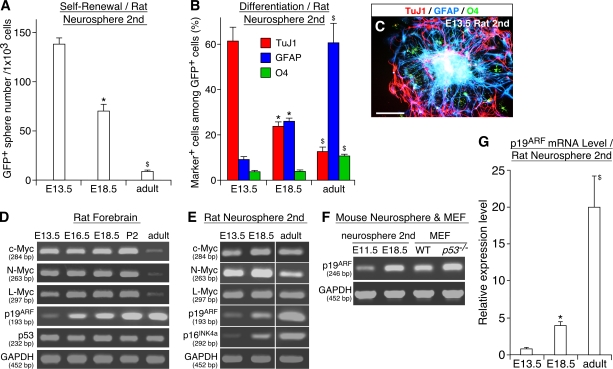
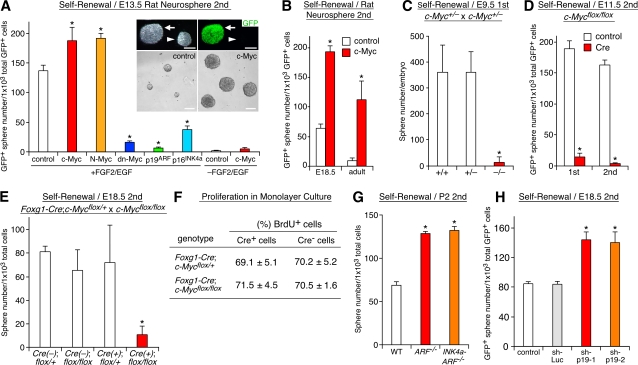
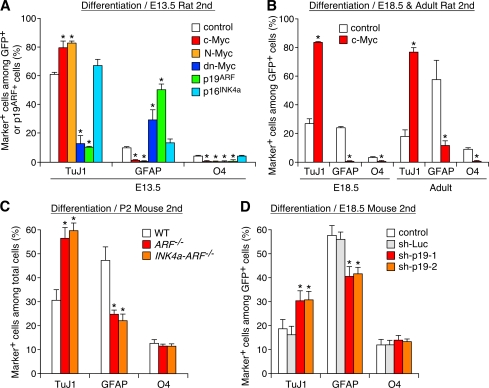
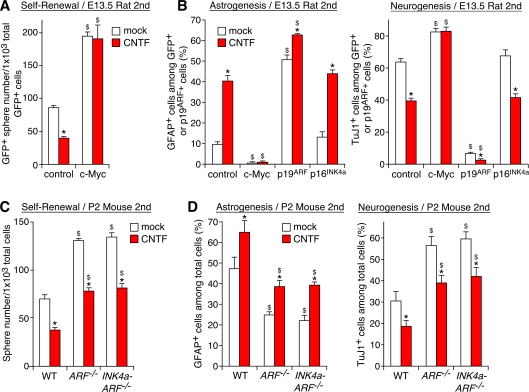
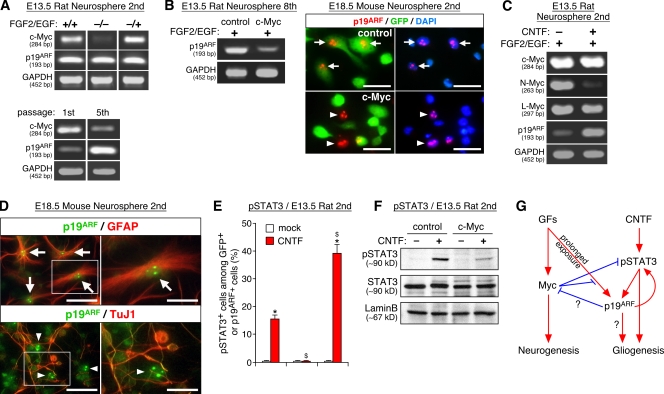
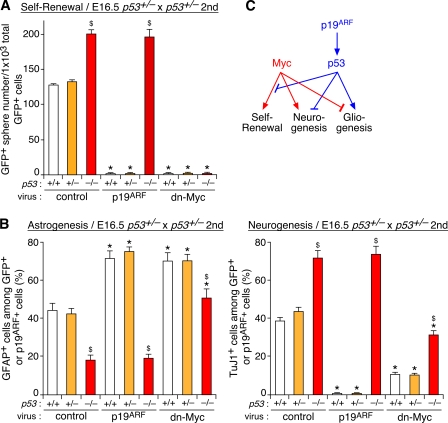
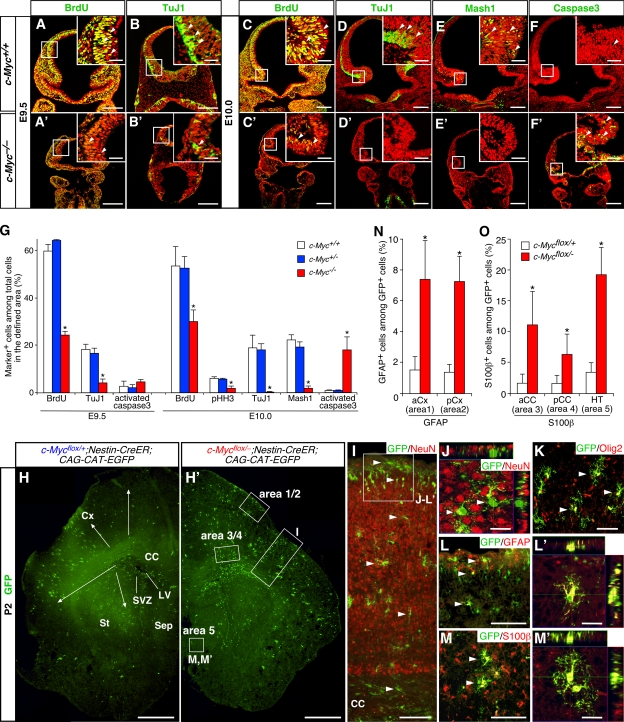
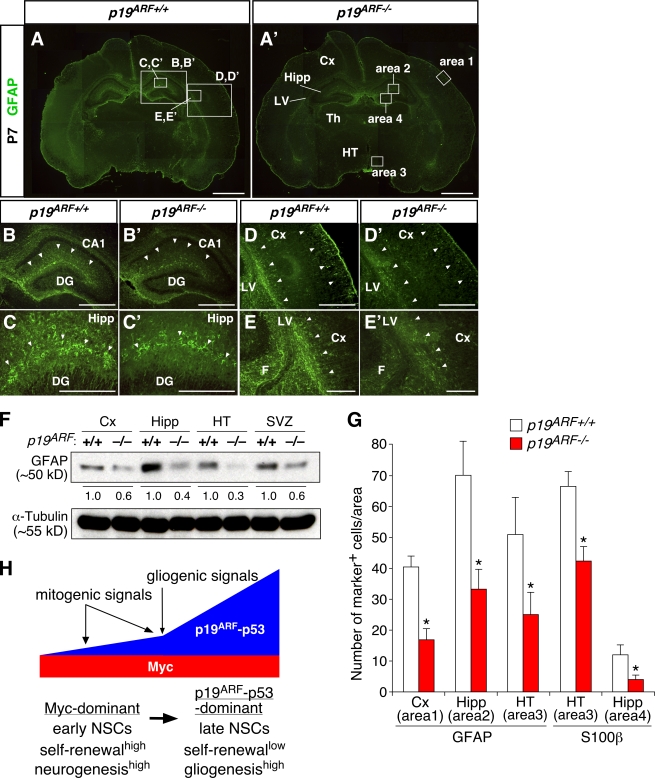
The modes of proliferation and differentiation of neural stem cells (NSCs) are coordinately controlled during development, but the underlying mechanisms remain largely unknown. In this study, we show that the protooncoprotein Myc and the tumor suppressor p19(ARF) regulate both NSC self-renewal and their neuronal and glial fate in a developmental stage-dependent manner. Early-stage NSCs have low p19(ARF) expression and retain a high self-renewal and neurogenic capacity, whereas late-stage NSCs with higher p19(ARF) expression possess a lower self-renewal capacity and predominantly generate glia. Overexpression of Myc or inactivation of p19(ARF) reverts the properties of late-stage NSCs to those of early-stage cells. Conversely, inactivation of Myc or forced p19(ARF) expression attenuates self-renewal and induces precocious gliogenesis through modulation of the responsiveness to gliogenic signals. These actions of p19(ARF) in NSCs are mainly mediated by p53. We propose that opposing actions of Myc and the p19(ARF)-p53 pathway have important functions in coordinated developmental control of self-renewal and cell fate choices in NSCs.
Figures
References
-
- Bachoo, R.M., E.A. Maher, K.L. Ligon, N.E. Sharpless, S.S. Chan, M.J. You, Y. Tang, J. DeFrances, E. Stover, R. Weissleder, et al. 2002. Epidermal growth factor receptor and Ink4a/Arf: convergent mechanisms governing terminal differentiation and transformation along the neural stem cell to astrocyte axis. Cancer Cell. 1:269–277. - PubMed
-
- Barnabe-Heider, F., J.A. Wasylnka, K.J. Fernandes, C. Porsche, M. Sendtner, D.R. Kaplan, and F.D. Miller. 2005. Evidence that embryonic neurons regulate the onset of cortical gliogenesis via cardiotrophin-1. Neuron. 48:253–265. - PubMed
-
- Berns, K., E.M. Hijmans, and R. Bernards. 1997. Repression of c-Myc responsive genes in cycling cells causes G1 arrest through reduction of cyclin E/CDK2 kinase activity. Oncogene. 15:1347–1356. - PubMed
-
- Bruggeman, S.W., and M. van Lohuizen. 2006. Controlling stem cell proliferation: CKIs at work. Cell Cycle. 5:1281–1285. - PubMed
-
- Bruggeman, S.W., M.E. Valk-Lingbeek, P.P. van der Stoop, J.J. Jacobs, K. Kieboom, E. Tanger, D. Hulsman, C. Leung, Y. Arsenijevic, S. Marino, and M. van Lohuizen. 2005. Ink4a and Arf differentially affect cell proliferation and neural stem cell self-renewal in Bmi1-deficient mice. Genes Dev. 19:1438–1443. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
